Here are three short videos that discuss helpful nutritional supplements for those who have taken the COVID vaccine. They help to mitigate the harm caused by the endless overproduction of the spike protein which results in constant inflammation, damage to the immune system, and other adverse effects. God bless.
(January 10, 2022, Prophet John Leary) Jesus Christ:“Trust in Me because My power is greater than the evil ones. You are seeing more sickness coming over your people, but the Covid vaccines and boosters will not help you. They will only reduce your immune system, and they could make you die. Trust in your supplements of vitamin C, D3 2000, elderberry extract, and Hawthorn. Your natural immunity is much stronger than the Pharmaceutical companies’ false vaccines. All the Covid vaccines are designed to kill people eventually, and produce the spike proteins that cause Covid to spread, especially with the omicron variant. Put your trust in My words against the demon’s lies and the evil people’s lies.”
A peer-reviewed medical study shows two, common, over-the-counter supplements, combine to destroy the spike protein of SARS-CoV-2. This article reprints the peer-reviewed study and tells you what supplements THEY used that wiped-out the spike protein. Maybe, people who took the “vaccines” can use this to wipe out the spike proteins inside themselves, that are making many of them sick, and killing many others?
In the interest of full disclosure, I do NOT sell any of the supplements mentioned here and earn NO MONEY from anyone, for passing along this information to you. I do this as a public service.
This peer-reviewed paper was published in March of 2021, but no one in the media bothered to tell the public. The only thing the media did was push the “vax.” Now, a lot of people are dead, dying, or very sick from the vax.
It seems that the messenger RNA in the vax, causes our human cells to “express a spike protein” like the one on the Coronavirus that causes COVID. Except human cells are not supposed to “express a spike protein.”
Below, is the study which showed two over-the-counter food supplements, Bromelain and Acetylcysteine (NAC), when used together – not separately – cause the spike protein bindings to fall apart and dissolve into nothing. Images of the results are below!
The Combination of Bromelain and Acetylcysteine (BromAc) Synergistically Inactivates SARS-CoV-2 by
1 Department of Surgery, St. George Hospital, Sydney, NSW 2217, Australia 2 Mucpharm Pty Ltd., Sydney, NSW 2217, Australia 3 CIRI, Centre International de Recherche en Infectiologie, Team VirPatH, Univ Lyon, Inserm, U1111, Université Claude Bernard Lyon 1, CNRS, UMR5308, ENS de Lyon, F-69007 Lyon, France 4 Hospices Civils de Lyon, EMR 3738 (CICLY), Lyon 1 Université, F-69921 Lyon, France 5 St. George & Sutherland Clinical School, University of New South Wales, Sydney, NSW 2217, Australia 6 Laboratoire de Virologie, Institut des Agents Infectieux (IAI), Hospices Civils de Lyon, Groupement Hospitalier Nord, F-69004 Lyon, France * Author to whom correspondence should be addressed. † These authors contributed equally to this work. ‡ These authors contributed equally to this work. Viruses 2021, 13(3), 425; https://doi.org/10.3390/v13030425 Received: 31 January 2021 / Revised: 25 February 2021 / Accepted: 1 March 2021 / Published: 6 March 2021 (This article belongs to the Special Issue Vaccines and Therapeutics against Coronaviruses)
Introduction The recently emergent severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the causative agent of coronavirus disease 2019 (COVID-19), which can range from asymptomatic to severe and lethal forms with a systemic inflammatory response syndrome. As of 21 February 2021, over 111 million confirmed cases have been reported, with an estimated overall mortality of 2.2% [1]. There are currently few therapeutic agents proven to be beneficial in reducing early- and late-stage disease progression [2]. While there are fortunately many vaccine candidates, their widespread availability for vaccination may not be immediate, the length of immune protection may be limited [3,4], and the efficacy of the vaccines may be reduced by novel SARS-CoV-2 variants. The continued exploration of effective treatments is therefore still needed.
Structurally, SARS-CoV-2 contains surface spike proteins, membrane proteins, and envelope proteins, as well as internal nucleoproteins that package the RNA. The spike protein is a homotrimer glycoprotein complex with different roles accomplished through dynamic conformational modifications, based in part on disulfide bonds [5]. It allows the infection of target cells by binding to the human angiotensin-converting enzyme (ACE2) receptors, among others, which triggers proteolysis by transmembrane protease serine 2 (TMPRSS2), furin, and perhaps other proteases, leading to virion and host cell membrane fusion [6,7].
The entry of viruses into mammalian cells, or “virus internalization”, is a key mechanism of enveloped virus infection and is based on dynamic conformational changes of their surface glycoproteins, namely, as mediated by disulfide bond reduction and regulated by cell surface oxydoreductases and proteases [5,8,9,10,11]. SARS-CoV-2 entry into host cells has been shown to start with destabilization of the spike protein through allosteric mechanical transition, which induces a conformational change from the closed “down” state to open “up” state of the receptor binding domain (RBD) of the spike protein [12,13]. The conformational changes of RBD and virus binding are induced by TMPRSS2 or Cathepsin L, which trigger the transition from the pre-fusion to post-fusion state [5,12,13]. The energy liberated by disulfide bond reduction increases protein flexibility, which is maximal when the reduced state is complete [8], thus allowing the fusion of host–virus membranes, which is otherwise impossible due to the repulsive hydration forces present before reduction [5].
Bromelain is extracted mainly from the stem of the pineapple plant (Ananas comosus) and contains a number of enzymes that give it the ability to hydrolyze glycosidic bonds in complex carbohydrates [14]. Previous studies have indicated that Bromelain removes the spike and hemagglutinin proteins of Semliki Forest virus, Sindbis virus, mouse gastrointestinal coronavirus, hemagglutinating encephalomyelitis virus, and H1N1 influenza viruses [15,16]. As a therapeutic molecule, it is used for debriding burns. Acetylcysteine is a powerful antioxidant that is commonly nebulized into the airways for mucus accumulation and is also used as a hepatoprotective agent in paracetamol overdose. Most importantly in the present context, Acetylcysteine reduces disulfide bonds [17]. Moreover, the association of the spike and envelope proteins by their respective triple cysteine motifs warrants the hypothesis of impacting virion stability following disulfide bridge disruption by the action of Acetylcysteine [18]. The combination of Bromelain and Acetylcysteine (BromAc) exhibits a synergistic mucolytic effect that is used in the treatment of mucinous tumors [19,20] and as a chemosensitizer of several anticancer drugs [21]. These different actions are due to the ability of BromAc to unfold the molecular structures of complex glycoproteins, thus allowing binding to occur because of the high affinity between RBD and ACE2.
Therefore, in the current study we set out to determine whether BromAc can disrupt the integrity of SARS-CoV-2 spike and envelope proteins and subsequently examine its inactivation potential against in vitro replication of two viral strains, including one with a spike mutant alteration of the novel S1/S2 cleavage site.
Materials and Methods 2.1. Materials
Bromelain API was manufactured by Mucpharm Pty Ltd (Kogarah, Australia) as a sterile powder. Acetylcysteine was purchased from Link Pharma (Cat# AUST R 170803; Warriewood, Australia). The recombinant SARS-COV-2 spike protein was obtained from SinoBiological (Cat# 40589-V08B1; Beijing, China). The recombinant envelope protein was obtained from MyBioSource (Cat# MBS8309649; San Diego, CA, USA). All other reagents were from Sigma Aldrich (St. Louis, MO, USA).
2.2. Recombinant Spike and Envelope Gel Electrophoresis
The spike or envelope proteins were reconstituted in sterile distilled water according to the manufacturer’s instructions, and aliquots were frozen at −20 °C. Two and a half micrograms of spike or envelope protein were incubated with 50 or 100 µg/mL Bromelain, 20 mg/mL Acetylcysteine, or a combination of both in Milli-Q water. The control contained no drugs. The total reaction volume was 15 µL each. After 30 min incubation at 37 °C, 5 µL of sample buffer was added into each reaction. A total of 20 µL of each reaction was electrophoresed on an SDS-PAGE (Cat# 456-1095; Bio-Rad Hercules, CA, USA). The gels were stained using Coomassie blue.
2.3. UV Spectral Detection of Disulfide Bonds in Spike and Envelope Proteins
The method of Iyer and Klee for the measurement of the rate of reduction of disulfide bonds has been used to detect disulfide bonds in spike and envelope proteins [22]. The recombinant SARS-CoV-2 spike protein at a concentration of 3.0 µg/mL in phosphate-buffered saline (PBS) (pH 7.0) containing 1 mM ethylenediaminetetraacetic acid (EDTA) was incubated with 0, 10, 20, 40, and 50 µL of Acetylcysteine (0.5 M), agitated at 37 °C for 30 min followed by equivalent addition of Dithiothreitol (DTT) (0.5 M), and agitated for a further 30 min at 37 °C. The spike protein was incubated in parallel only with DTT (0.5 M) as before without any Acetylcysteine and agitated at 37 °C for 30 min. The absorbance was then read at 310 nm. UV spectral detection of disulfide bonds in the envelope protein was performed in a similar manner.
2.4. SARS-CoV-2 Whole Virus Inactivation with BromAc
Fully respecting the World Health Organization (WHO) interim biosafety guidance related to the coronavirus disease, the SARS-CoV-2 whole virus inactivation tests were carried out with a wild-type (WT) strain representative of early circulating European viruses (GISAID accession number EPI_ISL_578176). A second SARS-CoV-2 strain (denoted as ∆S), reported through routine genomic surveillance in the Auvergne-Rhône-Alpes region of France, was added to the inactivation tests due to a rare mutation in the spike S1/S2 cleavage site and its culture availability in the laboratory (GISAID accession number EPI_ISL_578177).
2.5. Replication Kinetics by Real-Time Cell Analysis
To compare the in vitro replication capacity of both WT and ∆S SARS-CoV-2 strains, replication kinetics were determined by measuring the electrode impedance of microelectronic cell sensors on the xCELLigence Real-Time Cell Analyzer (RTCA) DP Instrument (ACEA Biosciences, Inc., San Diego, CA, USA). Vero cells were seeded at 20,000 cells per well on an E-Plate 16 (ACEA Biosciences, Inc., San Diego, CA, USA) and incubated with the same media conditions as described previously at 36 °C with 5% CO2. After 24 h, SARS-CoV-2 culture isolates were inoculated in triplicate at a multiplicity of infection of 10−2. Mock infections were performed in quadruplicate. Electronic impedance data (cell index) were continuously collected at 15-min intervals for 6 days. Area under the curve analysis of normalized cell index, established at time of inoculation, was then calculated at 12-h intervals. At each interval, cell viability was determined by normalizing against the corresponding cell control. Tukey multiple comparison tests were used to compare each condition on GraphPad Prism (software version 9.0; San Diego, CA, USA).
Results 3.1. Alteration of SARS-CoV-2 Spike and Envelope Proteins
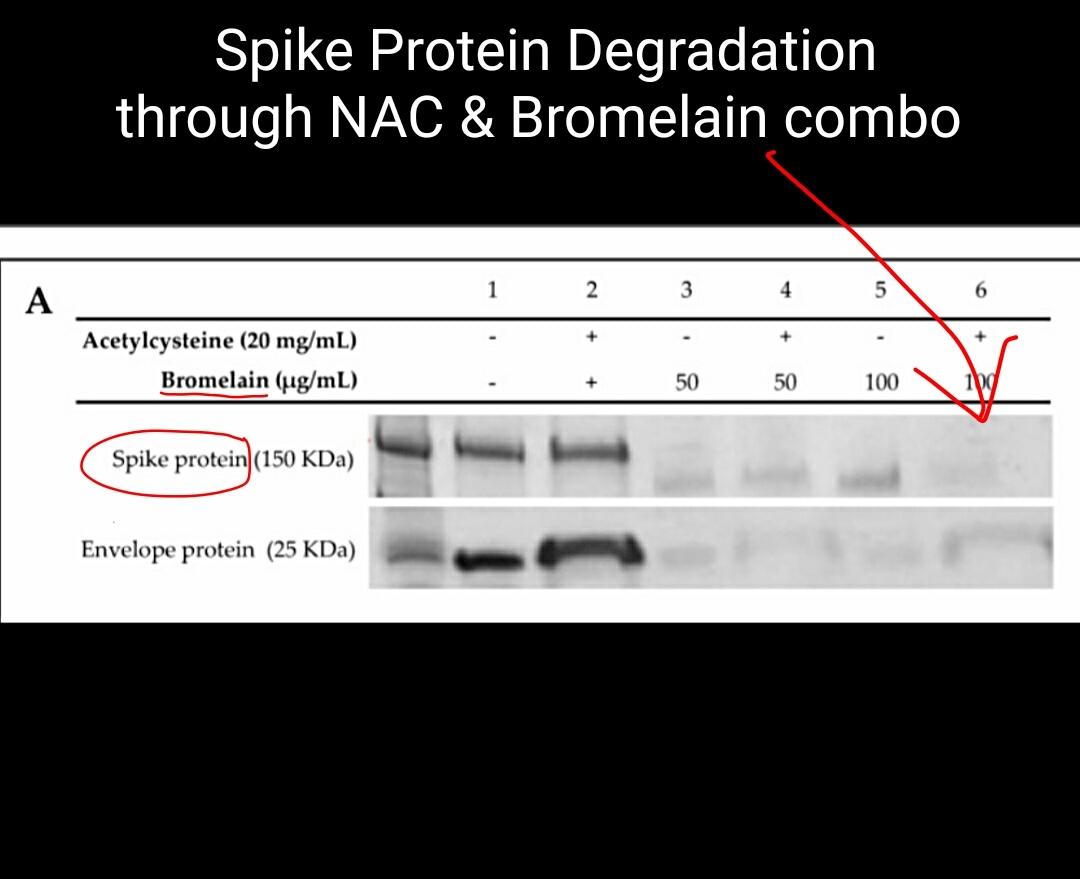
Treatment of the spike protein with Acetylcysteine alone did not show any alteration of the protein, whereas concentrations of Bromelain at 50 and 100 µg/mL and BromAc at 50 and 100 µg/20 mg/mL resulted in protein alteration (Figure 1A). Treatment with Acetylcysteine on the envelope protein did not alter the protein, whereas treatment with Bromelain at 50 and 100 µg/mL and BromAc at 50 and 100 µg/20 mg/mL also resulted in near complete and complete fragmentation, respectively (Figure 1A).
Figure 1. (A) Bromelain and Acetylcysteine present a synergistic effect on severe acute respiratory syndrome coronavirus (SARS-CoV-2) spike and envelope protein destabilization. SDS-PAGE of the recombinant SARS-CoV-2 spike protein S1 + S2 subunits (150 kDa) and envelope protein (25 kDa). Proteins were treated with 20 mg/mL Acetylcysteine alone, 100 and 50 µg/mL Bromelain alone, and a combination of 100 and 50 µg/20 mg/mL BromAc. (B) Disulfide reduction of recombinant SARS-CoV-2 spike protein by Acetylcysteine. The differential assay between Acetylcysteine (Ac) and Dithiothreitol (DTT) for the reduction of disulfide bonds found on the spike protein indicates that Acetylcysteine reduces 42% of the disulfide bonds before the addition of DTT. The remaining bonds are reduced by DTT to produce the chromogen detected at 310 nm. (C) Disulfide reduction of recombinant SARS-CoV-2 envelope protein by Acetylcysteine. The differential assay between Acetylcysteine (Ac) and Dithiothreitol (DTT) for the reduction of disulfide bonds found on the envelope protein indicates that Acetylcysteine reduces 40% of the bonds before the addition of DTT.
3.2. UV Spectral Detection Demonstrates the Alteration of Disulfide Bonds in Spike and Envelope Proteins
The comparative reduction of disulfide bonds on the spike protein between DTT alone and DTT with Acetylcysteine demonstrated a 42% difference (Figure 1B), based on the slope of the graphs [0.002599/0.006171 (100) = 42 %]. Acetylcysteine was thus able to reduce 58% of the disulfide linkages in the sample, after which the remaining disulfide bonds were reduced by DTT to produce the chromogen that was monitored in the spectra. Similarly, the differential assay between Acetylcysteine and DTT for the reduction of disulfide bonds found in the envelope protein [0.007866/0.01293 (100) = 60%] indicates that Acetylcysteine reduces 40% of the disulfide bonds before the addition of DTT (Figure 1C).
3.3. In Vitro SARS-CoV-2 Inactivating Potential of Bromelain, Acetylcysteine, and BromAc
For both SARS-CoV-2 strains tested, the untreated virus controls at 105.5 and 104.5 TCID50/mL yielded typical cytopathic effects (CPE), and no cytotoxicity was observed for any of the drug combinations on Vero cells. Optical CPE results were invariably confirmed by end-point Neutral Red cell staining. Overall, Bromelain and Acetylcysteine treatment alone showed no viral inhibition, all with CPE comparable to virus control wells, whereas BromAc combinations displayed virus inactivation in a concentration-dependent manner (Figure 2). Treatment on 104.5 TCID50/mL virus titers (Figure 2B,D) yielded more consistent inhibition of CPE for quadruplicates than on 105.5 TCID50/mL virus titers (Figure 2A,C).
Figure 2. Cell lysis assays demonstrated in vitro inactivation potential of Acetylcysteine and Bromelain combined (BromAc) against SARS-CoV-2. Cell viability was measured by cell staining with Neutral Red, where optical density (OD) is directly proportional to viable cells. Low OD would signify important cell lysis due to virus replication. The wild-type (WT) SARS-CoV-2 strain at 5.5 and 4.5 log10TCID50/mL titers (A and B, respectively) showed no inhibition of cytopathic effect (CPE) for single agent treatment, compared to the mock treatment virus control condition. BromAc combinations were able to inhibit CPE, compared to the mock infection cell controls. Treatment of a SARS-CoV-2 spike protein variant (∆S) with a mutation at the S1/S2 junction at 5.5 and 4.5 log10TCID50/mL titers (C and D, respectively) showed similar results. Bars represent the average of each quadruplicate per condition, illustrated by white circles. Ordinary one-way ANOVA was performed, using the mock treatment virus control as the control condition (**** p < 0.0001, *** p < 0.0005, ** p < 0.003, and * p < 0.05).
Based on the virus inactivation guidelines established by the WHO, a robust and reliable process of inactivation will be able to reduce replication by at least 4 logs [Log10 reduction value (LRV) = (RT-PCR Ct treatment – RT-PCR Ct control)/3.3; as 1 log10 ≈ 3.3 Ct]. As such, RT-PCR was performed on the RNA extracts to directly measure virus replication. For the wild-type (WT) strain at 104.5 TCID50/mL, successful LRV > 4 were observed with 1 out of 4 wells, 2 out of 4 wells, 3 out of 4 wells, and 4 out of 4 wells for 25, 50, 100 and 250 µg/20 mg/mL BromAc, respectively (Figure 3). It is worth noting that at 105.5 TCID50/mL, LRV were slightly below the threshold at, on average, 3.3, with 3 out of 4 wells and 2 out of 4 wells for 100 and 250 µg/20 mg/mL BromAc, respectively (Table 1). For the spike protein mutant (∆S) at 104.5 TCID50/mL, no successful LRV > 4 was observed for 25 µg/20 mg/mL BromAc, but it was observed in 4 out of 4 wells for 50, 100, and 250 µg/20 mg/mL BromAc (Figure 3). Of note, at 105.5 TCID50/mL, LRV were slightly below the threshold at, on average, 3.2, with 1 out of 4 wells, 2 out of 4 wells, and 4 out of 4 wells for 50, 100, and 250 µg/20 mg/mL BromAc, respectively (Table 1). Overall, in vitro inactivation of both SARS-CoV-2 strains’ replication capacity was observed in a dose-dependent manner, most strongly demonstrated at 100 and 250 µg/20 mg/mL BromAc against 104.5 TCID50/mL of virus.
Figure 3. Threshold matrix of log10 reduction values (LRV) of in vitro virus replication 96 h after BromAc treatment on WT and ∆S SARS-CoV-2 strains at 5.5 and 4.5 log10TCID50/mL titers. LRV were calculated with the following formula: LRV = (RT-PCR Ct of treatment—RT-PCR Ct virus control)/3.3; as 1 log10 ≈ 3.3 Ct. The color gradient matrix displays the number of quadruplicates per condition yielding an LRV > 4, corresponding to a robust inactivation according to the WHO. WT = wild-type; ∆S = S1/S2 spike mutant.
Table 1. Log10 reduction values (LRV) of in vitro virus replication 96 h after BromAc treatment on WT and ∆S SARS-CoV-2 strains at 5.5 and 4.5 log10TCID50/mL titers. LRV were calculated with the following formula: LRV = (RT-PCR Ct of treatment – RT-PCR Ct virus control)/3.3; as 1 log10 ≈ 3.3 Ct. Each replicate is described. TCID50/mL = Median Tissue Culture Infectious Dose; WT = wild-type; ∆S = S1/S2 spike mutant.
Real-time cell analysis demonstrated comparable replication kinetics for both WT and ∆S SARS-CoV-2 strains (Figure 4). No significant difference in cell viability was observed between WT and ∆S at any time point. From 48 h post-infection, WT and ∆S cell viability were significantly different compared to the mock infection (p < 0.05).
Figure 4. SARS-CoV-2 replication capacity of WT and ∆S SARS-CoV-2 measured by Real-Time Cell Analysis. Data points correspond to area under the curve analysis of normalized cell index (electronic impedance of RTCA established at time of inoculation) at 12-h intervals. Cell viability was then determined by normalizing against the corresponding cell control. WT = wild-type; ∆S = S1/S2 spike mutant.
Discussion The combination of Bromelain and Acetylcysteine, BromAc, synergistically inhibited the infectivity of two SARS-CoV-2 strains cultured on Vero cells. Protein confirmation and its molecular properties are dependent on its structural and geometric integrity, which are dependent on both the peptide linkages and disulfide bridges. Acetylcysteine, as a good reducing agent, tends to reduce the disulfide bridges and hence alter the molecular properties of most proteins. This property has been widely exploited in the development of several therapies (chronic obstructive pulmonary disease, allergic airways diseases, cystic fibrosis, pseudomyxoma peritonei, etc.) [20,23,24,25,26,27]. More recently, Acetylcysteine has been used in the development of therapies for respiratory infections such as influenza and COVID-19 [28,29,30], where the integrity of the spike protein is vital for infection [12,13]. A hypothesized mechanism of action could be the unfolding of the spike glycoprotein and the reduction of its disulfide bonds.
The SARS-CoV-2 spike protein is the cornerstone of virion binding to host cells and hence represents an ideal therapeutic target. A direct mechanical action against this spike protein is a different treatment strategy in comparison to most of the existing antiviral drugs, which prevents viral entry in host cells rather than targeting the replication machinery. BromAc acts as a biochemical agent to destroy complex glycoproteins. Bromelain’s multipotent enzymatic competencies, dominated by the ability to disrupt glycosidic linkages, usefully complement Acetylcysteine’s strong power to reduce disulfide bonds [17]. Amino acid sequence analysis of the SARS-CoV-2 spike glycoprotein identified several predetermined sites where BromAc could preferentially act, such as the S2’ site rich in disulfide bonds [31], together with three other disulfide bonds in RBD [32]. In parallel, the role of the glycosidic shield in covering the spike, which is prone to being removed by BromAc, has been highlighted as a stabilization element of RBD conformation transitions as well as a resistance mechanism to specific immune response [5,33,34].
Mammalian cells exhibit reductive functions at their surface that are capable of cleaving disulfide bonds, and the regulation of this thiol-disulfide balance has been proven to impact the internalization of different types of viruses, including SARS-CoV-2 [8,35,36,37,38]. Both ACE2 and spike proteins possess disulfide bonds. When all the spike protein RBD disulfide bonds were reduced to thiols, ACE2 receptor binding to spike protein became less favorable [8]. Interestingly, the reduction of ACE2 disulfide bonds also induced a decrease in binding [8]. Moreover, other reports suggested that Bromelain alone could inhibit SARS-CoV-2 infection in VeroE6 cells through an action on disulfide links [39,40]. As such, the loss of SARS-CoV-2 infectivity observed after pre-treatment with BromAc could be correlated to the cumulative unfolding of the spike and envelope proteins, with a significant reduction of their disulfide bonds by Acetylcysteine, demonstrated in vitro.
Interestingly, a similar effect of BromAc was observed against both WT and ∆S SARS-CoV-2. The main difference in amino acid sequences between SARS-CoV-2 and previous SARS-CoV is the inclusion of a furin cleavage site between S1 and S2 domains [41]. This distinct site of the spike protein and its role in host spill-over and virus fitness is a topic of much debate [41,42,43,44]. Of note, ∆S, which harbors a mutation in this novel S1/S2 cleavage site and alters the cleavage motif, exhibits no apparent difference in replication capacity compared to the WT strain. The slightly increased sensitivity of ∆S to BromAc treatment is therefore not due to a basal replication bias, but the mutation could perhaps be involved in enhancing the mechanism of action of BromAc. These results would nevertheless suggest that, from a threshold dose, BromAc could potentially be effective on spike mutant strains. This may be a clear advantage for BromAc over specific immunologic mechanisms of a spike-specific vaccination [3,4].
To date, different treatment strategies have been tested, but no molecules have demonstrated a clear antiviral effect. In addition, given the heterogeneous disease outcome of COVID-19 patients, the treatment strategy should combine several mechanisms of action and be adapted to the stage of the disease. Thus, treatment repurposing remains an ideal strategy against COVID-19, whilst waiting for sufficient vaccination coverage worldwide [45,46]. In particular, the development of early nasal-directed treatment prone to decreasing a patient’s infectivity and preventing the progression towards severe pulmonary forms is supported by a strong rationale. Hou et al. demonstrated that the first site of infection is the nasopharyngeal mucosa, with secondary movement to the lungs by aspiration [47]. Indeed, the pattern of infectivity of respiratory tract cells followed ACE2 receptor expression, decreasing from the upper respiratory tract to the alveolar tissue. The ratio for ACE2 was five-fold greater in the nose than in the distal respiratory tract [40]. Other repurposing treatments as a nasal antiseptic have been tested in vitro, such as Povidone-Iodine, which has shown activity against SARS-CoV-2 [48]. In the present study, we showed the in vitro therapeutic potential of BromAc against SARS-CoV-2 with a threshold efficient dose at 100 µg/20 mg/mL. As animal airway safety models in two species to date have exhibited no toxicity (unpublished data), the aim is to test nasal administration of the drug in a phase I clinical trial (ACTRN12620000788976). Such treatment could help mitigate mild infections and prevent infection of persons regularly in contact with the virus, such as health-care workers.
Although our results are encouraging, there are a number of points to consider regarding this demonstration. Namely, the in vitro conditions are fixed and could be different from in vivo. Any enzymatic reaction is influenced by the pH of the environment, and even more so when it concerns redox reactions such as disulfide bond reduction [9]. The nasal mucosal pH is, in physiological terms, between 5.5 and 6.5 and increases in rhinitis to 7.2–8.3 [49]. Advanced age, often encountered in SARS-CoV-2 symptomatic infections, also induces a nasal mucosa pH increase [49]. Such a range of variation, depending on modifications typically induced by a viral infection, may challenge the efficacy of our treatment strategy. Further in vitro experiments to test various conditions of pH are ongoing, but ultimately, only clinical studies will be able to assess this point. Our experiments were led on a monkey kidney cell line known to be highly permissive to SARS-CoV-2 infectivity. With the above hypothesis of S protein lysis thiol-disulfide balance disruption, BromAc efficacy on SARS-CoV-2 should not be influenced by the membrane protease pattern. Reproducing this experimental protocol with the human pulmonary epithelial Calu-3 cell line (ATCC® HTB-55™) would allow these points to be addressed, as virus entry is TMPRSS2-dependent and pH-independent, as in airway epithelium, while virus entry in Vero cells is Cathepsin L-dependent, and thus pH-dependent [50].
Overall, results obtained from the present study in conjunction with complementary studies on BromAc properties and SARS-CoV-2 characterization reveal a strong indication that BromAc can be developed into an effective therapeutic agent against SARS-CoV-2.
Conclusions There is currently no suitable therapeutic treatment for early SARS-CoV-2 aimed at preventing disease progression. BromAc is under clinical development by the authors for mucinous cancers due to its ability to alter complex glycoprotein structures. The potential of BromAc on SARS-CoV-2 spike and envelope proteins stabilized by disulfide bonds was examined and found to induce the unfolding of recombinant spike and envelope proteins by reducing disulfide stabilizer bridges. BromAc also showed an inhibitory effect on wild-type and spike mutant SARS-CoV-2 by inactivation of its replication capacity in vitro. Hence, BromAc may be an effective therapeutic agent for early SARS-CoV-2 infection, despite mutations, and even have potential as a prophylactic in people at high risk of infection.
Author Contributions
Conceptualization, J.A., K.P., S.J.V., and D.L.M.; methodology, J.A., G.Q., K.P., S.B., and A.H.M.; validation, J.A., G.Q., K.P., V.K., S.B., and A.H.M.; investigation, J.A., G.Q., K.P., V.K., S.B., and A.H.M.; writing—original draft preparation, G.Q., K.P., V.K, A.H.M., E.F., and S.J.V.; supervision, D.L.M. and E.F.; project administration, S.J.V.; funding acquisition, S.J.V. and D.L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research is partly funded by Mucpharm Pty Ltd., Australia.
Data Availability Statement
A preprint of this manuscript was archived on http://www.biorxiv.org (accessed on 31 January 2021) due to the emergency of COVID-19.
Conflicts of Interest
David L. Morris is the co-inventor and assignee of the Licence for this study and director of the spin-off sponsor company, Mucpharm Pty Ltd. Javed Akhter, Krishna Pillai, and Ahmed Mekkawy are employees of Mucpharm Pty Ltd. Sarah Valle is partly employed by Mucpharm for its cancer development and is supported by an Australian Government Research Training Program Scholarship. Vahan Kepenekian thanks the Foundation Nuovo Soldati for its fellowship and was partly sponsored for stipend by Mucpharm Pty Ltd.
References
1 John’s Hopkins University Coronavirus Resource Centre. COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). Available online: https://coronavirus.jhu.edu/map.html(accessed on 7 February 2021).
2 Song, Y.; Zhang, M.; Yin, L.; Wang, K.; Zhou, Y.; Zhou, M.; Lu, Y. COVID-19 treatment: Close to a cure?–a rapid review of pharmacotherapies for the novel coronavirus. J. Antimicrob. Agents2020, 56, 106080. [Google Scholar] [CrossRef] [PubMed]
3 Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet2020, 396, 479–488. [Google Scholar] [CrossRef]
4 Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet2020, 396, 467–478. [Google Scholar] [CrossRef]
6 Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res.2020, 176, 104742. [Google Scholar] [CrossRef]
7 Vankadari, N.; Wilce, J.A. Emerging WuHan (COVID-19) coronavirus: Glycan shield and structure prediction of spike glycoprotein and its interaction with human CD26. Microbes Infect.2020, 9, 601–604. [Google Scholar] [CrossRef]
8 Hati, S.; Bhattacharyya, S. Impact of Thiol-Disulfide Balance on the Binding of Covid-19 Spike Protein with Angiotensin-Converting Enzyme 2 Receptor. ACS Omega2020, 5, 16292–16298. [Google Scholar] [CrossRef] [PubMed]
9 Lavillette, D.; Barbouche, R.; Yao, Y.; Boson, B.; Cosset, F.L.; Jones, I.M.; Fenouillet, E. Significant redox insensitivity of the functions of the SARS-CoV spike glycoprotein: Comparison with HIV envelope. Biol. Chem.2006, 281, 9200–9204. [Google Scholar] [CrossRef][Green Version]
10 Mathys, L.; Balzarini, J. The role of cellular oxidoreductases in viral entry and virus infection-associated oxidative stress: Potential therapeutic applications. Opin. Ther. Targets2016, 20, 123–143. [Google Scholar] [CrossRef][Green Version]
11 Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science2020, 367, 1260–1263. [Google Scholar] [CrossRef][Green Version]
12 Moreira, R.A.; Guzman, H.V.; Boopathi, S.; Baker, J.L.; Poma, A.B. Quantitative determination of mechanical stability in the novel coronavirus spike protein. Nanoscale2020, 12, 16409–16413. [Google Scholar] [CrossRef]
13 Moreira, R.A.; Guzman, H.V.; Boopathi, S.; Baker, J.L.; Poma, A.B. Characterization of Structural and Energetic Differences between Conformations of the SARS-CoV-2 Spike Protein. Materials2020, 13, 5362. [Google Scholar] [CrossRef] [PubMed]
14 Amini, A.; Masoumi-Moghaddam, S.; Morris, D.L. Utility of Bromelain and N-Acetylcysteine in Treatment of Peritoneal Dissemination of Gastrointestinal Mucin-Producing Malignancies; Springer: New York, NY, USA, 2016. [Google Scholar]
15 Schlegel, A.; Schaller, J.; Jentsch, P.; Kempf, C. Semliki Forest virus core protein fragmentation: Its possible role in nucleocapsid disassembly. Rep.1993, 13, 333–347. [Google Scholar] [CrossRef]
16 Greig, A.S.; Bouillant, A.M. Binding effects of concanavalin A on a coronavirus. J. Comp. Med.1977, 41, 122–126. [Google Scholar]
17 Pillai, K.; Akhter, J.; Chua, T.C.; Morris, D.L. A formulation for in situ lysis of mucin secreted in pseudomyxoma peritonei. J. Cancer2014, 134, 478–486. [Google Scholar] [CrossRef]
19 Pillai, K.; Akhter, J.; Morris, D.L. Assessment of a novel mucolytic solution for dissolving mucus in pseudomyxoma peritonei: An ex vivo and in vitro study. Pleura Peritoneum2017, 2, 111–117. [Google Scholar] [CrossRef] [PubMed]
20 Valle, S.J.; Akhter, J.; Mekkawy, A.H.; Lodh, S.; Pillai, K.; Badar, S.; Glenn, D.; Power, M.; Liauw, W.; Morris, D.L. A novel treatment of bromelain and acetylcysteine (BromAc) in patients with peritoneal mucinous tumours: A phase I first in man study. J. Surg. Oncol.2021, 47, 115–122. [Google Scholar] [CrossRef] [PubMed]
21 Pillai, K.; Mekkawy, A.H.; Akhter, J.; Badar, S.; Dong, L.; Liu, A.I.; Morris, D.L. Enhancing the potency of chemotherapeutic agents by combination with bromelain and N-acetylcysteine—An in vitro study with pancreatic and hepatic cancer cells. J. Transl. Res.2020, 12, 7404–7419. [Google Scholar]
22 Iyer, K.S.; Klee, W.A. Direct spectrophotometric measurement of the rate of reduction of disulfide bonds. The reactivity of the disulfide bonds of bovine -lactalbumin. Biol. Chem.1973, 248, 707–710. [Google Scholar]
23 Zhang, Q.; Ju, Y.; Ma, Y.; Wang, T. N-acetylcysteine improves oxidative stress and inflammatory response in patients with community acquired pneumonia: A randomized controlled trial. Medicine2018, 97, 45. [Google Scholar] [CrossRef] [PubMed]
25 Calzetta, L.; Rogliani, P.; Facciolo, F.; Rinaldi, B.; Cazzola, M.; Matera, M.G. N-Acetylcysteine protects human bronchi by modulating the release of neurokinin A in an ex vivo model of COPD exacerbation. Biomed Pharm.2018, 103, 1–8. [Google Scholar] [CrossRef]
26 Cazzola, M.; Calzetta, L.; Facciolo, F.; Rogliani, P.; Matera, M.G. Pharmacological investigation on the anti-oxidant and anti-inflammatory activity of N-acetylcysteine in an ex vivo model of COPD exacerbation. Res.2017, 18, 26. [Google Scholar] [CrossRef] [PubMed]
28 Suhail, S.; Zajac, J.; Fossum, C.; Lowater, H.; McCracken, C.; Severson, N.; Laatsch, B.; Narkiewicz-Jodko, A.; Johnson, B.; Liebau, J. Role of Oxidative Stress on SARS-CoV (SARS) and SARS-CoV-2 (COVID-19) Infection: A Review. Protein J.2020, 39, 1–13. [Google Scholar] [CrossRef]
29 De Flora, S.; Balansky, R.; La Maestra, S. Rationale for the use of N-acetylcysteine in both prevention and adjuvant therapy of COVID-19. FASEB J.2020, 34, 13185–13193. [Google Scholar] [CrossRef]
30 Guerrero, C.A.; Acosta, O. Inflammatory and oxidative stress in rotavirus infection. World J. Virol.2016, 5, 38. [Google Scholar] [CrossRef] [PubMed]
31 Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell2020, 181, 281–292. e6. [Google Scholar] [CrossRef]
32 Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.K.; Huang, I.C.; Xu, K.; Vasilieva, N.; et al. Receptor and viral determinants of SARS-coronavirus adaptation to human ACE2. EMBO J.2005, 24, 1634–1643. [Google Scholar] [CrossRef] [PubMed][Green Version]
33 Watanabe, Y.; Allen, J.D.; Wrapp, D.; McLellan, J.S.; Crispin, M. Site-specific glycan analysis of the SARS-CoV-2 spike. Science2020, 369, 330–333. [Google Scholar] [CrossRef]
34 Casalino, L.; Gaieb, Z.; Goldsmith, J.A.; Hjorth, C.K.; Dommer, A.C.; Harbison, A.M.; Fogarty, C.A.; Barros, E.P.; Taylor, B.C.; McLellan, J.S. Beyond shielding: The roles of glycans in the SARS-CoV-2 spike protein. ACS Cent. Sci.2020, 6, 1722–1734. [Google Scholar] [CrossRef]
35 Ryser, H.; Levy, E.M.; Mandel, R.; DiSciullo, G.J. Inhibition of human immunodeficiency virus infection by agents that interfere with thiol-disulfide interchange upon virus-receptor interaction. Natl. Acad. Sci. USA1994, 91, 4559–4563. [Google Scholar] [CrossRef] [PubMed][Green Version]
36 Kennedy, S.I. The effect of enzymes on structural and biological properties of Semliki forest virus. Gen. Virol.1974, 23, 129–143. [Google Scholar] [CrossRef]
37 Schlegel, A.; Omar, A.; Jentsch, P.; Morell, A.; Kempf, C. Semliki Forest virus envelope proteins function as proton channels. Rep.1991, 11, 243–255. [Google Scholar] [CrossRef]
38 Compans, R.W. Location of the glycoprotein in the membrane of Sindbis virus. New Biol.1971, 229, 114–116. [Google Scholar] [CrossRef] [PubMed]
40 Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B. Tracking changes in SARS-CoV-2 Spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell2020, 182, 812–827. [Google Scholar] [CrossRef] [PubMed]
41 Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Biol.2020, 30, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
42 Jaimes, J.A.; Millet, J.K.; Whittaker, G.R. Proteolytic Cleavage of the SARS-CoV-2 Spike Protein and the Role of the Novel S1/S2 Site. iScience2020, 23, 101212. [Google Scholar]
44 Hoffmann, M.; Kleine-Weber, H.; Pohlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Cell2020, 78, 779–784. [Google Scholar] [CrossRef] [PubMed]
45 Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. Engl. J. Med.2020, 383, 2439–2450. [Google Scholar] [CrossRef]
46 Andersen, P.I.; Ianevski, A.; Lysvand, H.; Vitkauskiene, A.; Oksenych, V.; Bjoras, M.; Telling, K.; Lutsar, I.; Dumpis, U.; Irie, Y.; et al. Discovery and development of safe-in-man broad-spectrum antiviral agents. J. Infect. Dis.2020, 93, 268–276. [Google Scholar] [CrossRef]
Akhter J, Quéromès G, Pillai K, Kepenekian V, Badar S, Mekkawy AH, Frobert E, Valle SJ, Morris DL. The Combination of Bromelain and Acetylcysteine (BromAc) Synergistically Inactivates SARS-CoV-2. Viruses. 2021; 13(3):425. https://doi.org/10.3390/v13030425
Chicago/Turabian Style
Akhter, Javed, Grégory Quéromès, Krishna Pillai, Vahan Kepenekian, Samina Badar, Ahmed H. Mekkawy, Emilie Frobert, Sarah J. Valle, and David L. Morris. 2021. “The Combination of Bromelain and Acetylcysteine (BromAc) Synergistically Inactivates SARS-CoV-2” Viruses 13, no. 3: 425. https://doi.org/10.3390/v13030425
I, “a soul”, recently rediscovered an important video that had been worked on last year, but I had not gotten the opportunity to place it on this blog, MaryRefugeOfSouls. The video is about the Covid-19 vaccines and following the Church Magisterium. It is based on a message given by Our Lord, Jesus Christ, in April 2021. So, the video has been updated and finally finished. And I am very pleased to be finally presenting it to you all for your consideration.
Both this video and my video on healing those with the Covid-19 vaccines can only be found on this blog, MaryRefugeOfSouls, and my Brighteon video channel. Unfortunately, I cannot place them on YouTube due to censorship issues. However, if you are ever searching for a video created by me, “a soul”, you can find them all on my Brighteon channel. https://www.brighteon.com/channels/1asoul
Finally, I am including in this blog post, the PDF documents for both videos. This is for those who prefer to read the content instead of watching the videos. It is also for those who want to translate the materials.
Firstly, thank you to everyone for your prayers for my health. After five hospital and two skilled nursing facility stays these past nine months since January 2022, I am expecting to go home very soon. I know that all of your prayers have blessed me and I am so grateful.
Secondly, thank you to everyone who has sent me prayer requests. I am not able to respond to every email, but I hold everyone’s intentions close to my heart. God bless you all and your loved ones.
Thirdly, as there are many newcomers to this blog, MaryRefugeOfSouls, I want to re-share some previous content that has been featured on the blog before I got sick. As the newcomers may not know that on this prophecy blog, I “a soul”, have provided much information that is relevant to many timely topics. Below are important blog links and independent news sources that I follow on a regular basis. The links are broken down into the following topics:
Covid-19 vaccines (important information)
Credible independent news sources
Nuclear war (how to prepare for staying safe)
The Warning (Illumination of Conscience)
Moment of Death
Lastly, during these past nine months that I have been sick, I have received a few emails from blog followers who are generously volunteering to shelter and board conservative priests who are in crisis for their traditional faith (mainly due to their love of Jesus in the Holy Eucharist). The Blessed Mother requested that I maintain a list for the coming time of persecution in the USA and several families have already volunteered. Please know that I am still maintaining the list, but the original spreadsheet is on my broken computer, which has not been updated since I got sick. However, once I return home and my brother is finished with building a new computer, I will be able to work on this project again. Meanwhile, please forgive me for any lack of acknowledgment, as I am setting aside these special emails for when I can return to doing this project. Your generosity has been noted by me and I thank you very much.
Thank you, dearly beloved ones. May God bless abundantly everyone who follows this blog, MaryRefugeOfSouls. I love you all.
Jesus said: “My people, you are seeing more signs of the end times with earthquakes, volcanoes going off, and possible food shortages leading to a famine. Now you are seeing another sign of cars leaving their homes for a refuge in your vision. This means the Warning would have to come first along with a mini-judgment, and the conversion time. This is why I have been warning people to have their backpacks ready to put in their cars, when your angel will lead you to the nearest refuge of protection. I told you that I would bring My Warning before the next deadly virus is released, or when a major life threatening event is about to happen. I am always watching out to protect My faithful. You also need to have your soul prepared with frequent Confession at least monthly. The more you are prepared, the more you can warn others to be prepared for the coming tribulation. Trust in Me to warn you with My inner locution when it is time to come to My refuges.”
Monday, October 25, 2021
Jesus said: “My people, you are in the end times and you are close to coming to My refuges, and eventually you will see My New Earth and New Heavens as in the vision of the New Jerusalem. I call My people to repent of their sins and have their souls prepared for My judgment. Only those souls, who live holy and faithful lives, will be allowed through the gates of My New Jerusalem. I will give all sinners one last chance to be saved when I will bring My Warning. You will see your life review and a judgment of where you are in life. You will see how you have a choice to convert your life to be with Me. Otherwise, the people, who ignore Me and reject Me, will be locked out of heaven, and they will enter the gates of hell’s flames. Repent of your sins now while you can, and you can be protected in My refuges from all of the evil events that are coming. Trust in Me and My angels to protect you from the evil ones.”
(Mass intention for Sr. Faith) Jesus said: “My people, you may not have come to Sr. Faith’s funeral, but she is more than thankful for having this Mass offered up for her. You just saw a movie on purgatory, and you appreciate more how these souls need your prayers and Masses to help them get up to heaven. Sr. Faith was a beautiful person of faith, and she is happy to be with Me now. Give thanks to Me for giving her life to you.”
Sunday, October 24, 2021
Jesus said: “My people, in today’s Gospel I healed Bartimeus of his blindness, because he had a deep faith that I could heal him. This deep faith in My healing power, is what is needed in all of My healing. In the same way, those people, who are sick with the Covid virus, or those, who are sick from the Covid shots, can be healed if they have faith that I can heal them. The vaccinated people can also be healed using the Good Friday oil or the exorcism water. I am showing you this angel, because your guardian angels will be leading you to My refuges after the Warning. This is because the deadly virus will be coming once you are in the safety of My refuges. Once the deadly virus is released, you will be seeing many bodies on the ground who will be dying from a bleeding virus. Nobody will want to remove these bodies for fear of getting the deadly virus. All the vaccinated people will have destroyed immune systems that will not protect them from the new virus. Only the vaccinated people, who are at My refuges or healed by the Good Friday oil or the exorcism water, will survive. The new vaccines will kill even more people because these new shots will be more deadly than the Covid shots, so refuse any new vaccines. My faithful will need to come to My refuges to be healed by looking on My luminous cross. Trust in My angel protection for healing and providing for your needs.”
Saturday, October 23, 2021
(St. John of Capistrano) Jesus said: “My people, the parable about the barren fig tree is a lesson to My faithful that I expect all of you to bear fruit in your missions. You can bear fruit by doing good deeds for your neighbor, but the best fruit is when you can bring souls to Me by conversion. Be a good example of My love and prayer so others can imitate your love for Me. I desire to save all souls, but they must come to Me of their own free will. Another fruit is that you can come to Confession often, and repent of your sins. I love all of you as My children, and I do not want to lose even one soul. Pray for the conversion of poor sinners among your intentions. I pray that My children will be open to take My words to heart and act on them.”
Jesus said: “My son, you know how cold it gets in your area and how important it is to keep warm if you did not have access to your natural gas. This is why I gave you a vision of the wood you have available. You have roughly enough wood for about four months and kerosene for a month. For your house I could multiply this fuel and you could continue to heat your house for the winters of the tribulation time. You also have some propane that I could multiply for cooking and baking your bread. You have been spoiled when you can use natural gas all of these years, but it is much more work to heat your house with other fuels. Trust in Me to provide heat for your refuge house and also for the other dwelling I will provide for the rest of your people.”
Jesus said: “My son, I have been talking to you about being prepared for a possibility of a large tsunami wave that could come from a landside of a volcano into the Atlantic Ocean. I am confirming that the one world people are planning to use the HAARP machine to cause earthquakes and volcanic activity on the Canary Islands. This will continue the present activity which could cause a landslide into the Atlantic Ocean. This would precipitate a large tsunami that could kill millions of our people on the East Coast. I will raise My hand to prevent the killing of millions of people by the one world people by warning you to get to high ground should this event occur.”
Friday, October 22, 2021
(St. John Paul II) Jesus said: “My people, in today’s Gospel (Luke 12:54-59) you have a sense of the coming end times. When you see the dark clouds, you know that a rain storm is upon you. If you can read the weather in the sky, why is it that you cannot recognize that you are living in the end time? A time is coming when the evil ones will try and force another shutdown on you when they will release the next deadly virus on the people. The signs of the end times can be seen in the growing lack of faith in Me, as fewer people are coming to Sunday Mass. Very few people are coming to repent of their sins in Confession. You are seeing signs of a coming famine, as you are seeing some empty shelves in the grocery stores. Volcanoes are going off in various places, and people are getting sick from the flu and the Covid virus variants. I have had My refuge builders put up their refuges for when the tribulation will come, so My faithful will have a safe haven. My angels will protect you from the Antichrist, the evil people, and the demons. After My Warning, and the conversion time of six weeks, I will call My faithful to My refuges with My inner locution. When I call you, you need to grab your backpack and leave your homes within twenty minutes for My refuges. Trust in Me to protect you at My refuges.”
Jesus said: “My son, I told you to keep a watch on the volcanic eruptions going on in the Canary Islands. You have read how the earthquakes are up to 4.8 from 4.5 in the previous days. The lava flow is continuing and the activity has gotten worse. If there is a landside into the ocean, it will be important to warn the people of any possible tsunami as soon as possible, so people can be warned to get to higher ground away from the coast. If a large tsunami hits America, it could cause major devastation and drown millions of people. Pray that you may be warned enough in advance to avoid any drowning if a large tsunami comes. Trust in Me to protect all of My refuges along the East Coast.”
Thursday, October 21, 2021
(Alice Sutherland intention) Jesus said: “My people, the division I spoke of is how families would be divided over who believes in Me, and those who do not believe in Me. Everyone is called to Me in the Gospel, but not everyone wants to give up their life to Me, your Creator. You have many other divisions that are being created by your government and your health officials. Biden is pushing to vaccinate everyone with the poisonous Covid shots, so you have those who are vaccinated against those who do not want to take these shots that will be a slow death. The one world people are using lies with the media to reduce the population with these Covid shots. You do not need these shots because your natural immunity is better. Biden is forcing people to choose between the shots or their jobs. It is better to refuse these shots and trust in finding another means to support yourselves. Those people, who are vaccinated, can be healed with the Good Friday oil, the exorcism water, or by calling on Me to heal them. All means of healing occur when you believe that I can heal you. The most important division is who will follow My laws and repent, or who will refuse My love and refuse to repent. My followers will be rewarded in heaven, but the non-believers will be cast into hell forever. Choose life with Me for those people who want to be with Me forever in heaven.”
Alice: Jesus said: “My people, Alice was a stalwart and strong in the faith to keep the Christ the King shrine going over all these years. She is with Me in heaven for all that she suffered in life. Give thanks to Me for her beautiful life before you.”
Jesus said: “My people, when you want your teeth cleaned, you come to the dental hygienist. When you want your soul cleansed, you come to the priest in Confession. My sacrament of Reconciliation was instituted at the Last Supper, and it is to help sinners to cleanse their sins, and bring My sanctifying grace to heal their souls. In order to enter heaven, you need to have clean souls, and you need to repent of your sins as well. I died on the cross so I could bring salvation to all souls who seek My forgiveness. Trust in Me that I will offer all souls one last chance to repent of their sins at My Warning and in the conversion time.”
Jesus said:“My son, you have gone to this FEMA detention center in Indianapolis, Indiana and you could see the windows covered up and the vents for the gas chambers. There were crematoria inside to burn the bodies. These are places that the evil ones will be exterminating Christians and those people who do not go along with the one world people’s plans for a new world order. I have warned My faithful that I will call you to My refuges when the evil ones want to force the mark of the beast on everyone. My angels will protect My faithful at My refuges, even though some will be martyred.”
Jesus said: “My people, I have been calling on My people to push back on the Biden vaccine mandates. You are seeing some states passing laws against the vaccine mandates because they are unconstitutional. I call on My people to also stop this insanity of forcing the poisonous Covid shots on the young children. Continue also to keep protesting against the abortions of the unborn babies at the Planned Parenthood clinics, as you do on Saturday morning. The death culture people are now using deadly viruses and vaccines to kill many people in time. They also continue to kill the unborn babies in abortion and the elderly with euthanasia. By fighting these killers, you will be rewarded in heaven.”
Jesus said: “My people, you are seeing how important it is to store your three months of food for when you could see empty shelves in your stores. The famine is coming, and not only are you seeing inflation with higher prices for your food, but you are even beginning to see empty shelves because of your food shortages. I have told you before, if you do not heed My words of storing three months of food, you could be starving to death with closed empty stores. Trust in Me that I will multiply your food even now, if you ask Me and believe that I can do this.”
Jesus said: “My people, you are seeing attempts to move more containers off the ships at your docks by forcing people to work the docks 24 hours every day. One of the bottlenecks has been the union dock workers who have refused to accept more automated unloading of the ships. In other countries with automation, they unload the containers off the ships twice as fast as your dock workers without automated devices. Pray that your food can be distributed as soon as possible, but there will be shortages as you enter the coming famine. Trust in My multiplication of food for your survival.”
Jesus said: “My people, I have warned you not to take any Covid shots, and you will also have to reject taking the mark of the beast, or you could lose your soul to the Antichrist. In the Book of Revelation you are told not to take the mark of the beast and not to worship the Antichrist, or you could be lost in hell. (Rev. 14:9-11) Protect your family from taking this mark as well. At My Warning you can help your family to convert any wayward souls. Be patient and come to My refuges of protection with My angels protecting you from the evil ones.”
Jesus said: “My people, many of My faithful will soon be seeing more persecutions and even some of My people will face martyrdom. Your government is pushing hard to force the poisonous Covid vaccines on you. Some people are losing their jobs by not taking these shots. They will seek out the unvaccinated people and put you in detention death camps if they can catch you. When your lives are threatened, I will call you to the safety of My refuges. The refuges and My angels will protect you from the evil ones who will be trying to kill you. Trust in My protection throughout the whole tribulation, when My angels will put an invisible shield over you at My refuges and over My people on the way to My refuges.”
Wednesday, October 20, 2021
(St. Paul of the Cross) Jesus said: “My people, be prepared in your souls because you do not know the day of the Son of Man’s return to the earth at the judgment. The best preparation for your soul is frequent Confession and daily prayer as at daily Mass. The physical preparation for the coming tribulation is to be ready to leave for My refuges with your backpacks or roller boards ready to load into your car. My refuge builders are finishing up with last minute needs, or My angels will finish any needed preparations. Much will be expected of those people who know of the end times, and more will be expected from My refuge builders. Pray that you are doing all that I have asked of you. Trust in My protection and feeding you at My refuges.”
Jesus said: “My people, I have told you that the Covid shots are full of toxic graphene oxide that causes blood clots and it will kill people. Now you are reading how the evil leaders want to force the toxic Covid shots on your young children who rarely get sick from the Covid virus. The young children, who rarely get sick from the Covid virus, have more glutathione which is an antioxidant which helps keep the blood from clotting. Pray that your people will rise up against these toxic shots for the young children. The evil ones who want to reduce the population are forcing toxic Covid shots on the elderly, the workers, and now the young children. These evil ones should have a millstone put around their necks and be thrown into the sea for trying to kill My little ones. Pray to protect your children from these poisonous shots that the evil ones are trying to force on them.”
MESSAGE FROM OUR SAVIOR, JESUS CHRIST “SEEK YE FIRST THE KINGDOM OF GOD, AND ALL HIS RIGHTEOUSNESS” OCTOBER 18, 2021, MONDAY @ 4:18 P.M.
Anna Marie: My Lord, are you calling me?
Jesus: Yes my little one, I your Savior Jesus of Nazareth am calling you.
Anna Marie: My Lord may I ask please? Will you bow down and adore God your Eternal Holy Merciful Father who is the Alpha and Omega, the Creator of all life, of all that is visible and invisible?
Jesus: Yes little one, I your Divine Savior will now and will always bow down and adore my Holy Eternal Merciful Father who is the Alpha and Omega, the Creator of all life, of all that is visible and invisible.
Anna Marie: Please speak my Holy Lord, for your sinful servant is now listening. [A private message was given to Anna Marie.]
Jesus: My dear one, I have come to give you an important message for all my cherished Apostles. Please let them know that during these difficult days of persecution (Mother Mary appeared), I will be at their side. For anyone who is demanded to get the covid vaccine but chooses not to, I will help him or her and their family too. A bully only gets his way if he can intimidate another. With you, it will not be so. Stay strong and defend you rights as a human being created in God’s Holy Image and you will stand with others who are also being persecuted. I tell you now, your crown in Heaven will be great amongst the Saints and Angels because you endured my WILL, instead of giving in to those who rebuke me, their Savior. Stay strong, be mindful of my dearest Mother who is always there to comfort you during your trials and sufferings.
Jesus:BEHOLD, my Father’s Hand will strike your land because of the grievous evil your citizens continue to perpetrate against the innocence: the babies who are aborted, the exploitation of their body parts, and the little ones being used in human sacrifices to the enemy of God. His reign is nearly over where he and all his servants will be chained in hell and suffering unspeakable sufferings. Any soul who refuses to serve me, serves the devil and will be locked into the pit of hell.
Anna Marie: My dear Lord, may I ask you please? When will this wrath happen? Is it to take place during 2021?
Jesus: Yes, it shall be done this year.
Anna Marie: My dearest Lord, can we all pray to mitigate it?
Jesus: Yes, you can pray for its mitigation.
Anna Marie: Jesus, what should it be called?
Jesus: The Wrath of My Father.
Anna Marie: Jesus may I place it on a blessed Green Scapular for Mother Mary to help us pray for its mitigation?
Jesus: Yes, this should be done.
Anna Marie: My Lord, have mercy on your obedient but sinful Apostles. We are trying to remain faithful, but the anti-Pope is in control and we are all devastated in our daily lives. We cannot run to our Churches anymore. Many Priests tell us to get the vaccination and do not understand how trafficking witches are attacking your children! They don’t realize that major corporations are cursing their products targeting children and when parents buy these items, the children are terrified because most products have demons attached to them. Priests do not understand this, they are not listening to the Holy Spirit! Where do we go for help my Dearest Lord?
Jesus:As long as the words of Consecration have not been changed, your Masses are valid. You must trust that I AM with you, leading you. My Mother is praying with you and protecting you. You must at a minimum recite your holy Rosary and Chaplet of Divine Mercy daily. Otherwise, you may be consumed by evil in every aspect of your daily lives.
Jesus: My dear Apostles, very soon the Illumination of your Conscience will take place. At that time, your Priests will understand what will be required of them and so will you. You will need to make a sincere, heartfelt Sacramental Confession to cleans your soul. A WAR is on the horizon, prepare little ones, prepare now. Time is short and your opportunity to prepare is almost over.
Jesus: Now my little ones, pray! Pray all day long if possible. Seek me in Mass, seek me in Eucharistic Adoration. Seek me in all you do. Remember the Scriptures: “Seek ye first the Kingdom of God, and all His Righteousness” (St. Matthew 6:33).
Jesus: My dear one, ensure this message is posted quickly.
Anna Marie: Yes my Divine Savior, I will.
Jesus: Go my dear one, continue your chores and do as I have asked.
Anna Marie: Yes my Lord, I will. I love you Jesus and all your Apostles around the world love you dear Sweet Jesus!
Jesus: I love them all too. Your Divine Savior, Jesus Christ, Son of the Living God. (Mother Mary left after the message was completed.)
Once again, I (Maureen) see a Great Flame that I have come to know as the Heart of God the Father. He says:
“Children, do not be distracted by the errors and rumors surrounding this pandemic. All of this is just Satan’s way of confusing and distracting hearts and leading them away from trust. I am still in charge of every present moment. Therefore, do not allow your hearts to be casualties of Satan’s propaganda. Trust always in My Will for you. Place this trust in the cocoon of the Holy Mother’s Heart* where falsehoods cannot break in to destroy your peace.”
“After all this passes and the pandemic is history, you will look back and see more clearly all of Satan’s ploys and designs in this pandemic. The Ancient Enemy uses whatever is at hand to promote chaos.”
Read Psalm 3:3-4+
But thou, O Lord, art a shield about me, my glory, and the lifter of my head. I cry aloud to the Lord, and he answers me from his holy mountain.
+ Scripture verses asked to be read by God the Father. (Please note: all Scripture given by Heaven refers to the Bible used by the visionary. Ignatius Press – Holy Bible – Revised Standard Version – Second Catholic Edition.)
* The Immaculate Heart of the Blessed Virgin Mary.
Once again, I (Maureen) see a Great Flame that I have come to know as the Heart of God the Father. He says:
“These days, people are very careful about protecting themselves from the COVID virus, and rightly so. They see clearly what they should do to protect themselves and what they should avoid. But, while they are cautious about protecting their mortal lives, they disregard any caution in protecting their immortal life. They do not take note of that which weakens their relationship with Me. They do not try to protect their faith from marauders. They expose their faith to every kind of ‘germ’ which would challenge its well-being. They gather in large crowds of unbelievers without My Protection.”
“My children, your immortal life is what you make of it while on earth. When you respect My Commandments* and try to live in a way that pleases Me, it is as though you are wearing a mask which protects your soul. Make this a priority in your everyday existence as much as you do protection from disease.”
Read Colossians 3:1-4+
If then you have been raised with Christ, seek the things that are above, where Christ is, seated at the right hand of God. Set your minds on things that are above, not on things that are on earth. For you have died, and your life is hid with Christ in God. When Christ who is our life appears, then you also will appear with him in glory.
+ Scripture verses asked to be read by God the Father. (Please note: all Scripture given by Heaven refers to the Bible used by the visionary. Ignatius Press – Holy Bible – Revised Standard Version – Second Catholic Edition.)
* To LISTEN or READ the nuances & depth of the Ten Commandments given by God the Father from June 24 – July 3, 2021, please click here: https://www.holylove.org/ten/
Tuesday, October 5, 2021 (St. Faustina, Bl. Francis Seelos)
Jesus said: “My son, you see many mistakes in life made by many people, because you all have some imperfections in carrying out your work. It is hard to criticize others when you all have made mistakes in your own life. Let people correct their mistakes, and move on with their lives. You are seeing My glory in heaven in your vision. I am thankful for all of St. Faustina’s work to help all of you to understand a little about My Divine Mercy. You are faithful in praying your own Divine Mercy Chaplet at the 3:00 o’clock hour every day. I forgive all sinners of their sins in Confession when they repent of their sins. This is how forgiving I am that I have mercy on all repentant sinners. I even have given more graces to people who pray in front of My Divine Mercy picture. Give thanks and praise to Me for all that I do for My people.”
Monday, October 4, 2021 (St. Francis of Assisi)
Jesus said: “My people, I called Jonah to go to the people of Nineveh to warn them of My impending destruction of their city. Jonah fled away because he did not want to help his enemy. The evil from Nineveh was so blatant that My wrath was about to come down on them. I wanted to give these people one more chance by sending My reluctant prophet Jonah to warn them. After being thrown into the sea and being in a big fish for three days and three nights, I had the fish spew Jonah onto the shore. So Jonah went about Nineveh proclaiming that in forty days, their city would be destroyed. The people heeded this warning, and they stopped their evil ways, and they put on sackcloth and sat in the ashes. Because of their change, I relented of My intended destruction, and I did not carry it out. I see an even worse evil emanating from America, and I am calling My prophets in America to warn your people of a similar destruction that will fall on your people, unless you repent and change your evil ways. It appears that My warnings have fallen on deaf ears, and your reluctance to change your evil abortions and sexual sins is calling out for My justice. I am warning My faithful to be ready to leave for My refuges, because soon I will give you My inner locution that it is time to come to My refuges. Your destruction will be far worse than your Twin Tower incident on 9-11-01, so be prepared. Those people, who come to My refuges when I call them, will be protected from My coming wrath. Those people, who do not repent and do not seek My forgiveness, will suffer through the coming hell on earth. Trust in Me that in the end, the evil ones will be cast into hell, and My faithful will be brought into My Era of Peace.”
Jesus said: “My son, I have shown you how the souls suffer from the flames of hell, and there is no hope to escape hell through the iron bars. I am warning the souls on earth to be prepared for My coming Warning with frequent Confession. All sinners will be shown their life review and judgment. After the Warning there will be a time of conversion without any evil influence. This will be the last chance for sinners to repent and ask for My forgiveness of their sins. You will then be placed back into your bodies and in time so you will have a chance to change your lives. Those people, who do not repent, will suffer a hell on earth from the plagues and the Antichrist, and then forever in hell. Try to convert your relatives during the conversion time, because you do not want anyone to be lost in hell. Only converted believers will be allowed into My refuges, where My angels will protect them from the evil ones. Trust in My protection, and have no fear during the tribulation time.”
Jesus’ blessing goes out to Joanne for this Mass.
Sunday, October 3, 2021 (Right to Life Mass by Bishop Matano)
Jesus said: “My people, I am happy to see a Mass for Right to Life. Once a new conception of a human being occurs, there are two possibilities for the baby: either a live birth, or an aborted dead baby. At conception I put life into that first fertilized cell, and a guardian angel is assigned to that baby. Most of the babies survive the abortionist’s knife, and after birth they are allowed to fulfill My intended mission for that soul. In cases of abortion the baby has a shortened life, and the guardian angel assists bringing that soul to heaven as a martyr. I see all of the abortions performed in each country, and I am planning the severity of My justice on the percentage of abortions in each country. America has decisions and laws that make abortion legal to kill babies. I have mentioned before that your country will be taken over by the communists as a punishment for your abortions. Before the lives of My faithful are threatened, I will bring My Warning and a chance for conversion of souls. After the conversion time, I will call My believers to My angel protection at My refuges. I call on My faithful to pray and protest your abortions and your abortion laws. Keep praying at the abortion clinics to try and save My babies.”
Saturday, October 2, 2021 (Guardian angel day, Mark as my angel)
St. Mark said: “I am Mark and I stand before God as I was appointed to watch over you, my son. You could be a little more reverent and I urge you to read the Bible more. You are much easier to watch over because you are aware of all of the evil around you, thanks to the words of Jesus that you receive every day. I watch over you when you are driving in your car. Even when you had your car accidents, I was there guiding you. You are sensing the plans of the evil ones who want to give all of the Covid shot resisters trouble for not going along with the one world people’s plan. You also have prepared your refuge well. The Lord has shown you all of the other angels who are protecting your refuge. This will be your place of protection throughout the coming tribulation. You are blessed to be one of our Lord’s leaders to help protect the faithful who will come to your refuge. You have been shown the plans of the angels who will help build your high rise building to house the 5,000 people who will come. I will be helping you also to direct the people to safety. Just as you read how the angels protected the Israelites from their enemies, so your refuge angels will be protecting you with shields against any evil ones who want to attack you. Trust in our help in guiding you, protecting you, and providing for your daily Holy Communion. We will assist you also in the multiplication of your food, water, and fuels. Pray with us and give glory and thanks to God for all He does for you.”
Jesus said: “My people, your people are being distracted by your sports games, while the Democrat socialists are stealing your freedoms with vaccine mandates, and arbitrary Executive Orders from Biden. The vaccines are toxic with graphene oxide, and people will die with two or three shots. The media and Biden have lied to the people that vaccines are safe, and will protect people from the Covid virus. These are lies and the vaccines could kill you over time, and the vaccinated people still get sick. Now there is a war against the unvaccinated people and they are being turned into second class citizens with less rights than the illegal immigrants who are crossing the border illegally. There is discrimination with the vaccine mandates when Congress and the illegal immigrants are exempt from being forced to take the Covid shots. These shots are a means of reducing the population, and they are being forced on My faithful. Refuse to take these shots, even if they threaten to kill you. This is why I will soon bring My Warning before any attempts for martial law or a shutdown. After My Warning and the conversion time, I will call My faithful to My refuges, and I will bring My justice down on the evil ones who will be cast down into hell. Trust in My angel protection as I will defeat the evil ones at the Battle of Armageddon.”
Friday, October 1, 2021
(Rosemarie Thummler Funeral Mass) Rosemarie said: “I want to thank my family and friends for coming to my funeral. I am at peace and I will be in heaven shortly with my Lord. My church and my Lord were the center of my life. I was laughing with all of you when you mentioned how I loved desserts. I am praying for all of my family members and I ask for your prayers and one more Mass. I will be watching over you, and keep me in your loving hearts as I love all of you. I await the day when I can see all of you in heaven.” P.S. I have no more worries.
(St. Therese of Lisieux) Jesus said: “My people, when I told you the angels would be protecting your house from harm, I am showing you how your windows would not even break. Just as if they were made out of guerrilla glass, your windows will be protected. My angels are also protecting you from fire, bullets, bombs, or any evil people. You have seen a few miracles to show you how My angel protection is real, even if you cannot see My angels protecting you. I have given you a mission to set up a refuge at your house, and you have followed all of My suggestions. Now you have your solar batteries fixed and your solar panels are in place. Your off grid solar system will power your water pump and your two sump pumps in your basement, especially in the winter. You have beds, food, water from your well, and the fuels as I recommended. Be ready, for My refuge time is coming soon to protect you from the evil ones.”
Thursday, September 30, 2021 (St. Jerome)
Jesus said: “My people, I came on the earth to fulfill the words of the law in the Scriptures. It is important to know and understand My words, so you can live your life in the Light of My grace. That is why you should listen closely to the words of the Gospel that are read to you every day at Mass. This is why I wanted you to have your Magnificat or other books so you could remember My words and be able to recite them to others, if needed. You also have My prophets of today as there were prophets in the Old Testament. You need to listen to their words as well, because I send them to you just as I sent My disciples out to the towns to prepare the towns for when I would speak. So when My prophets or messengers go before you, they need to have a place to stay and eat, for they are worthy of their keep. Listen to My words that go before you, because you may not have this grace of My words for long. A time of the evil tribulation of the Antichrist is about to come before you, when I will call My faithful to My refuges for your protection. Trust in Me and My words of warning for that time.”
Jesus said: “My people, you are being told lies upon lies from the father of lies in Satan. It is a lie that you need a poisonous shot, because those people, who had the Covid virus, do not need a vaccine. The vaccines will kill you in a few years, but the media is lying to you that you need this shot. The media lies to you about all of the vaccinated people who are getting the Covid virus, and some are dying, or are deathly sick with blood clots. The real truth is that the vaccines and boosters are designed to kill you and reduce the population. The evil ones want to kill all those people, who refuse these shots, because you are against their deadly plans. Trust in Me that I will protect you at My refuges.”
Jesus said: “My people, you will be seeing lock downs coming to America as most of your sick and dying people are the vaccinated people. The more shots and boosters people take, the quicker they could develop symptoms and die. Refuse to take any Covid shot, boosters, or flu shots. They will all destroy your immune system, and you could die from the next corona virus if you take the shots. As the evil ones try to restrict your travel and freedoms, I will soon call you to My refuges of protection.”
Jesus said: “My people, your new governor in New York is becoming more dictatorial than your previous governor in trying to force everyone to get the Covid shots. You are seeing a shortage of workers in your hospitals and other critical jobs. Your governor is planning to bring in National Guard troops to fill any needed vacancies. This is against your separation of civilians from the military. She will use these same troops for the coming checkpoints to limit where you can travel. These are more signs of how close your refuge time is coming. Trust in Me and My angels to protect My faithful.”
Jesus said: “My people, your fuels could be in danger from the people who want to stop using your fossil fuels. Some people are trying to get others to use electric cars, but they do not realize this puts a strain on your power plants that are already at maximum capacity. When your fuels get scarce because of poor management of your fossil fuels, you may not have enough fuel for your cars. This will be enhanced if you have another virus shutdown. Have your backpacks or roller boards and refuges ready for these coming trials.”
Jesus said: “My people, the people who are running your government are radicals and they are against using fossil fuels. There are not enough means for supplying the energy you need with green sources. More restrictions on your fuels will cause the shortages that will restrict your travel. Trust in Me to multiply your fuels at your refuges.”
Jesus said: “My refuge builders need to fix up their refuges and their food supplies so they are ready to feed all the people who I will send to each refuge. You have heating and cooking needs that will require My multiplication of your wood, kerosene, propane and other fuels for your winter season that is coming. You will need these fuels when your natural gas lines go empty. You will also need propane and wood for cooking your food. You will have to adapt to refuge life without electricity and your natural gas. Trust in Me that My angels will assist you and I will multiply your fuels, food, and water.”
Jesus said: “My people, when the evil ones try to kill My faithful, I will bring My Warning to protect them, and I will bring My justice down on the evil ones. Your guardian angels will be leading My faithful to My refuges with the flames. Your refuges will be protected by My angels, and only believers with a cross on the forehead will be allowed into My refuges. This is why you need to try and convert your family members to the faith during the conversion time after the Warning. You will be so thankful for all of My refuges that will have invisible shields that will protect you from the evil ones. Trust in Me to multiply your food, water, and fuels that you will need for your survival.”
Wednesday, September 29, 2021 (St. Michael, St. Gabriel, St. Raphael)
St. Michael said: “I am Michael, and I stand before God. You are witnessing all the evil in the world that the Lord is allowing because of the free will of man. Toward the end of the tribulation there will be a great battle on earth called the Battle of Armageddon. I will be leading this battle with God’s angels and his faithful against the demons and the evil people. Just as I was victorious with God’s grace in casting the demons into hell out of heaven, so with God’s grace I will defeat the demons and the evil ones as they will again be cast into hell as well. Have no fear of the evil ones because I will be protecting the faithful at the Lord’s refuges.”
Jesus said: “My people, you are seeing the evil ones using your Covid trial to try and control you with their evil poisonous shots. They are threatening the unvaccinated with losing their jobs if they do not get vaccinated. Your people are stressed about their jobs, but they are not pushing back about these communist tactics. The Covid shots are not a total cure from Covid virus because some vaccinated people are still getting sick, and some are with worse complications than the Covid virus. Be aware that the vaccines are being used to reduce the population on purpose, and the vaccinated people are getting blood clots from the shots. When the next wild corona virus comes, the vaccinated people will be dying in the streets as I showed you in visions because their immune system is destroyed by the vaccines. Your leaders are taking away your freedoms, but they will pay when My justice falls on them. Be ready for My coming Warning, and after the Warning, you will be called to My refuges of protection. After the tribulation of the Antichrist you will see My angels will defeat the demons and the evil people at the Battle of Armageddon, and the evil ones will be cast into hell. Then I will renew the earth and bring My faithful into My Era of Peace.”
Jesus said: “My people, the unvaccinated people are under attack, and today you are seeing the workers in the health field being forced to take the poisonous Covid shots, or they would lose their job. This is an attack on your freedoms when you allow these threats against your jobs to force people to take these shots. The people who work at larger work places are allowed to get tested for Covid virus, but soon this option will be taken away for all workers, and they will need to take the shots also. You have seen some demonstrations and protests, but this has not stopped your President or your governor. These shots will kill people in a few years, and they are not really required of those people who have had the Covid virus. You are seeing total control from your evil masters. This will put more people out of work who refuse the shots. You already are having a worker shortage. Your people will either have to revolt, or come to My refuges to avoid this communist control. I will send My Warning when the evil ones try to kill or imprison My faithful. After My Warning, you will be called to My refuges. Be prepared.”
Monday, September 27, 2021 (St. Vincent de Paul)
Jesus said: “My people, I want My people to imitate St. Vincent de Paul in how he helped the poor. The more you help others instead of keeping things just for yourself, the more treasure you will be saving up in heaven for your judgment. You need to give your time to help others, your donations of money, and your prayers for converting sinners. You need to show your love for Me and your neighbor by helping others whenever you can. Do not be spiritually lazy, but do something for the poor and your family with your daily prayers and actions.”
Jesus said:“My people, souls in lower purgatory are those who were barely saved from hell. These are the lukewarm souls who had relatives praying for their souls to be saved. The souls in the flames need to have a minimum time before Masses and prayers could move them up out of the flames. All the prayers and Masses will be applied to them once they have done this minimum time. My faithful need to pray for your relatives who may still be in purgatory. You can also pray for the souls in purgatory who have no one to pray for them. You pray for people on earth, but do not forget to pray for the poor souls in purgatory so they can come to heaven in time.”
Sunday, September 26, 2021 (Mass for Camille Remacle,Carol’s dad)
Camille said: “Hello again, John. I am happy to talk with you again. In this vision you are seeing a great divide, and not between the vaccinated and unvaccinated, but between those who believe in God, and those who do not believe in God. Your mission of encouraging the believers to come to God’s refuges is truly about dividing the believers from the non-believers in God. At the refuges it is true that God’s angels will be protecting the believers from being harmed from the demons, or the evil people who are trying to kill the believers. After the coming Warning, God will be calling His believers to His refuges. During the conversion time, you need to reach out to evangelize Vic and all of the lukewarm souls in your families. Only the believers will be given the cross on the forehead that will enable them to enter God’s refuges. Keep praying for all of the souls who are away from God that this conversion time will be their means of being saved in God’s kingdom. I look forward to the day when you all can come to heaven.”
Saturday, September 25, 2021
Jesus said: “My son, you are preparing for your skype conference with your friends at Gospa Prayer house. In your vision I was showing you a river of living water from the Holy Spirit who will help speak through you the words of My messages. Call on the Holy Spirit to start your talk, and pray the St. Michael prayer to protect you from any evil ones who want to disrupt your presentation. In the first reading at Mass you heard of a wall of fire being put around Jerusalem to protect the people. My son, at your refuge I have shown you in the picture how your refuge angel, St. Meridia, is putting an invisible shield around your refuge so the evil ones will not see it during the tribulation. I have also shown you all of the tall angels standing side by side protecting all of My refuges on the whole perimeter of each refuge, even now. Give thanks and praise to Me for all of My protection, and My multiplication of your needs at the proper time.”
Jesus said: “My people, your hurricane season is not over and you are seeing one more strong storm that could hit your coast. You have had some severe damage this year, but your people are still far away from Me. You are being tested by your storms, and the Democrats who are causing major damage to your country. Your border being open has been a major disaster, but nothing is being done to stop the endless flow of illegal immigrants. If the Democrats stay in power, your country will soon fall apart with their disastrous policies. Pray for your people to fight the forced vaccine mandates. The illegal immigrants are not being vaccinated, so your Covid virus control is getting out of hand. You will be seeing a continuing disaster with your government spending and out of control deficits.”
Friday, September 24, 2021
Jesus said: “My people, in today’s Gospel I asked My disciples first, ‘Who do the people say I am?’ They replied: ‘Some say St. John the Baptist or one of the prophets. Then I asked them: ‘But who do you say that I am?’ St. Peter said: ‘Thou art the Christ, the Son of the living God.’ I told My apostles not to reveal this to anyone, but truly I am the Second Person of God in the Blessed Trinity. My people of today are fortunate to have My Blessed Sacrament that you receive in every Mass. I am truly Present in My Eucharist, and you have Me in your soul for a short time. This is a little taste of heaven on earth. I ask that you receive Me worthily in the state of grace without mortal sin, or you commit a sin of sacrilege if you are in mortal sin. I love all of My people so much, and I give you graces and your guardian angel to protect you from the temptations of the devil. I will protect you further at My refuges, where many angels are standing side by side all around the perimeter of a refuge’s property. This is the protection you will need from the Antichrist and the demons during the tribulation. Rejoice that I am protecting My refuges even now, just as you saw your friend in Louisiana when her refuge buildings were protected even from Hurricane Ida’s winds.”
Jesus said: “My people, your satellites are linked in to your banking system that moves money over the internet. You are seeing explosions in space because you have rogue satellites armed with missiles that could take down your communication satellites. This would be a threat to your banking system and your military defenses. On the ground you could see another means of attack on your electric grid from various hackers. If your enemy nations can take down your electric grid with hackers, you would have many people without money that would not move in your banks. The one world people will use hackers first. If that is unsuccessful, then they would use an EMP attack on your grid. Without electricity your country would be paralyzed. If such an event happens, I will call my people to My refuges. If your lives are threatened, I will bring My Warning, and then you will be called to My refuges. Be ready to come to My refuges, because some serious events will be going on.”
Jesus said: “My people, you are seeing many serious events about to commence, and I am warning your people to come to frequent Confession, and have your three months of food on hand. If your finances or your electric grid collapse, you will need to come to the protection of My refuges. You will be sharing My recent messages at your talk tomorrow, and warn the people not to take the poisonous Covid shots or the flu shots.”
Thursday, September 23, 2021 (St. Padre Pio)
Jesus said: “My people, the Temple was in ruins in Jerusalem, so I encouraged the people through Ezra, the prophet, to get lumber and stone to rebuild My Temple which was the center for their worship of Me. Today, I call on all of My believers to build up My Church by sharing your faith with others and evangelizing them to the faith. Fewer people are coming to worship Me, and you are more concerned with the temporary things of this world, instead of taking care of your soul in your spiritual life. Your soul is eternal and I created you to be with Me in heaven of your own free will. You need to confess your sins frequently in Confession to keep a pure soul ready to greet Me at your judgment when you die.”
St. Padre Pio said: “I thank you for remembering me as I suffered from the stigmata for many years. Offer up your pains and discomforts to the Lord for saving souls. Be a good example of faith to all of those people around you. My son, continue to wear your cross outside.”
Jesus said: “My people, you have been faithfully protesting outside of a local hospital so they would stop the Covid vaccine mandates for the nurses. This week the health care people have to make a choice between having a job or mandated Covid vaccine shots. Some people have families and they need the money to support their families. These are your front line health people that will be harmed from these shots. About 20% of the people taking these shots have complications. Your governor is threatening to bring in foreign health people. If many of your unvaccinated health people refuse these shots, you could see some severe shortages in your hospitals. My justice will fall heavy on Biden and those people threatening the people’s jobs. You may see more sick people from these shots. You can bless the vaccinated people with the Good Friday oil or they can drink the exorcism water.”
Jesus said: “My people, you still have more time for hurricanes to develop. You also are seeing volcanic eruptions in the Canary Islands that could threaten a tsunami with another major earthquake. You are seeing a lot of flooding in the East with heavy rainstorms. You are seeing some serious droughts and fires out West. These weather disasters are testing your people and they are a punishment for your abortions and your sexual sins.”
Jesus said: “My people, you are fortunate to have a working water well, because finding fresh water is important for your survival. This shortage of fresh water could get worse if you see your electricity shut down by a number of causes. At My refuges I will multiply your water because you need water for survival. This is why I have encouraged My refuge builders to have a supply of fresh water for the people.”
Jesus said: “My people, you remember several times that your lights did not work at night when your electricity stopped in a wide area power outage. I have given you some estimates that nearly 90% of your people would die of starvation if your electricity was stopped for a long time. This is why I have encouraged people to have at least three months supply of food in case your stores had empty shelves. Solar power and generators would be helpful in a long power outage. Call on Me if you are in need of food, and I will multiply what you have.”
Jesus said: “My people, if your authorities threaten the lives of the unvaccinated, I will call them to My refuges for My angel protection. When protection is needed, My angels will put a shield of invisibility over My faithful. At My refuges I will multiply your food, water, and fuels for your survival. Not only are the evil ones trying to kill people with these poisonous shots, but they want to kill all the people who are going against the leftist communism that is being forced on your people. I am having My refuge builders make refuges because this is how I will protect My faithful throughout the tribulation of the Antichrist. When I call you to My refuges with My inner locution, remember to leave your homes within 20 minutes with your backpack. Your guardian angels will lead you with a flame to the nearest refuge.”
Jesus said: “My people, you need to have no fear of the evil ones at your refuges because My angels will do battle for you against the demons and the evil people. My refuge builders have been stocking up some food, water, and fuels for what you will need during the tribulation. You cannot store enough supplies to last for less than 3 ½ years for all of the people who I will send to you. This is why I will have to multiply your needs just as I did for the people of Moses’s time in the desert. So trust in Me that My angels will protect you, and you will have enough supplies to survive. You must believe that I can do this for all of you.”
Jesus said: “My people, I told you that I would be with you always, even to the end of time. This is why My priests or My angels will supply you with daily Holy Communion. You will take a consecrated Host and place it in a monstrance at every refuge. This will allow you to be able to have perpetual Adoration. You will assign hours of Adoration for each person to have their time in front of My Blessed Sacrament. By having Me with you, you will have My grace to endure your time during the tribulation. Trust in My protection and My providing you with what you will need for your survival.”
Wednesday, September 22, 2021
Jesus said: “My people, in today’s Gospel you read how I breathed the Holy Spirit on My apostles so they were enabled to heal people’s diseases and to expel demons. My people of today have also received the Holy Spirit in your Baptism and Confirmation. This means you can pray over people in My Name, and they can be healed. You can also pray deliverance prayers as a group to expel demons, and when you pray, you can fast as well. Some people have addictions that are controlled by demons. By praying over such people in My Name you can expel the demon and heal the addiction. Some people are taking the Covid vaccine shots, and they have complications from the shots. You can pray over these people with My Good Friday oil or the exorcism water to heal their immune system. Trust in My power to heal people through My faithful as intercessors.”
Jesus said: “My son, your wife made an attempt to look for a bread machine and other flours for some variety. Take a look online to buy some of these things if you cannot find them in the store. It would be better to buy the 25 lb size bags of other flours if they are available. I am giving this message as a follow up to giving a better look online for what is available. Take some time to locate and make any purchases.”
Jesus said: “My people, you have been focused on the latest fraudulent election. This is a distraction from an ongoing pandemic virus that does not kill that many people. Now, you are seeing the one world people push nano particle vaccines that are making people sick or even killing some people. I told you before that you will next see another virus that will be much deadlier. You did not understand that the Chinese scientists were giving you the first part of a binary virus that is purposely designed to kill many people. This second virus will react with the vaccine and could kill many people as well. I will warn My people before this deadly virus is released. I will call My believers to My refuges where you will look upon My luminous cross, and you will be healed of any virus, even if you took the virus vaccine.”
The video has been transcribed below using the subtitles from the video. Slight spelling alterations (vaxxine-vaccine) and embellishment has been added for emphasis.
Father Savvas: “I would like to present to you a personal testimony of a Priest-Monk, who made the mistake of getting vaxxed. This is his confession. If anyone wants to know his name, I can tell you privately. He is a priest under Archbishop Ieronymos of Greece. I will be reading directly from the publication. Which fortunately, it is still on the internet.
[Priest-Monk]: “With a few words, I will relate my experience after taking my first dosage of the Pfizer vaccine.
God obstructed me with many signs before taking the vaccine, due to shortness in time I will not mention all of them.
As I was heading towards the vaccination center, right before I was getting into the queue I felt something was obstructing my approach. As I approached, I felt (smelled) a stench that surprised me.”
Father Savvas: “Said the monk. You see, God cautioned him and he still went ahead.”
[Priest-Monk]: “While I was taking the vaccine, others were waiting outside. As I was leaving I was unable to wear my Kalimafi (priest hat).
I felt a great shame within myself, and left holding my kalimafi in my hands.”
Father Savvas: “As you can see, he began to feel the demonic influence right away. As he was ashamed to wear his Kalimafi even though he is a priest.”
[Priest-Monk]: “Arriving home, I went to the bathroom to wash my face. Upon looking at the mirror, I was frightened at my face due to the expression I had.
The next day I went shopping at the supermarket and since it was still the period after Pascha (before the leave-taking), I would normally greet the shopkeepers with “Christos Anesti (Christ is Risen!)” or respond “Alithos Anesti (Indeed He is Risen)”.”
Father Savvas: “So as you know, for 40 days after the resurrection, we refrain from saying “Hello/Good day”, and replace the phrase with “Christ is risen”. And the other Christian should respond “Alithos Anesti” and not say “likewise”.
[Priest-Monk]: “As a Priest monk, I was surprised to discover that I was very ashamed to say “Christo Anesti” to the shopkeepers. This greatly overwhelmed me.”
Father Savvas: “He began to realize more intensely that something was not going well in his soul.”
[Priest-Monk]: “A day later I went and attended a divine service at a local church, but not to liturgize the service as the priest.
Upon entering the Altar I felt as if I was dead.”
Father Savvas: “You see a living spiritual person notices the difference straight away.”
[Priest-Monk]: “The joy I used to feel at the divine service was lost. It was as if I was not entering the altar of a Holy Church, but as if I had entered a room in a house. All these things surprised me, but at the time I did not believe they stemmed from the vax.
I saw familiar parishioners turn their faces away from me.
The next day I found out that my conscience was causing me terrible pain. It is as if I had been pierced in my heart with such pain that I had never felt before in my life.
I told this to a fellow Archimadrite (Monastic Priest), of what I was feeling, and he consoled me. He told me words along the lines of: ‘It’s nothing, don’t worry about it.’
Upon leaving, I found that this pain in my conscience was relentless and was deepening further within me. After that day forward I was in a deeply troubled state which lasted 13 days. I could not sleep or quiet down.
Now allow me to explain the most terrible part. Day and night I constantly saw Satan in front of me, his face 20mm distance from mine.
I went to sleep at night, and felt him hugging me, and I would get cold all over.
I would read the Salutations (Χαιρετισμούς) to the Virgin Mary and I would feel as if my blood was burning in my veins. I felt a foreign presence within me and it was judging me.
I felt a horror as if someone was saying: “You belong to me now.” “
Father Savvas: “Do you understand how terrible these things are? And very true because we personally know this person.
In turn, this proves another experience of another Priest-Monk who was doing exorcisms. And the demon being pressed told the truth while having a monologue: “Why am I telling you this? I don’t want to tell you this, but I am being pressed.”
The Priest-monk replied, “I am not pressing you.” The demon replied, “I am being forced to tell you.”
So the demon told him thus: “We did a ceremony at a Lodge in America for the vaccines.” The satanists performed a ceremony for the vaccines. Furthermore the demon said: “Those who take this vaccine will be unable to repent.” Now this may seem too harsh.
The Priest-monk then asked, “Why won’t they be able to repent?”
The demon responded, “Because I will be inside of them.”
You can see a correlation with the initial Priest-monk who was saying the same thing, that he felt Satan inside of him and saw him 20mm away from his face telling him, “you belong to me”.
The Priest-monk performing the exorcism, was having a dialogue with the demon. The demon was speaking through the demon-possessed person.
The demon told him: “Those who took the vaccine cannot repent because I am inside of them.” The Priest-monk asked, “How are you inside of them?”
The demon answered, “With the blood of the aborted fetuses.”
We have mentioned previously, that fetuses were used in the vaccine and were purposely murdered for their cells. These cells were extracted from the living fetuses by these atheist scientists and doctors who don’t hold anything sacred and holy.
They also remove the organs from a living fetus. If the fetus is dead, the organs and cells are useless. Therefore, they were not taking the fetuses from the waste bin. Which even if they were, does not make it morally right, as an abortion had taken place. However, in this case, these fetuses were specifically prepared for an abortion.
So the devil confesses, “I am already inside those who took it via the blood of the fetuses.”
So this confession of this demon, correlates with the Priest-monk who was seduced into taking the vaccine.
So as we previously read; he was saying he was ashamed to wear his kalimafi, to say ‘Christos Anesti’, how he felt dead whilst being in the holy altar, how everyone turned their faces away from him because his face was altered… how for 13 days he could not sleep or settle down, and most terrifying of all, day and night was seeing the face of Satan from his face at a 20mm distance continuously… and how he felt Satan hugging him, and though he was trying to read the Salutations, his blood was burning in his veins, and he felt someone saying to him, “You belong to me now.”
Thus the Priest-monk continues:
[Priest-Monk]: “I had stayed at my family home in case of an adverse reaction, after a few days I left.
At the Monastery where I currently reside, at the Divine Liturgy, I found that I could not understand a thing.
I felt as if I was dead, I was constantly rushing through the service, felt great anxiety, not a speck of joy, I felt as if I was not a priest or even a baptized Christian!
I reached a point unable to speak as if I had lost my voice. I felt my life was dark and a constant state of despair.”
Father Savvas: “You see this person made this mistake and is being humbled. This means he has an ecclesialogical conscience. This is very important.
As there are others who have made this mistake, and after seeing this mistake do not confess it, so that they may warn others.
Let’s continue reading.”
[Priest-Monk]: “While I was in this hopeless state, a familiar family came to visit the monastery. I spoke with the mother.
She said to me, ‘Father, why are you speaking like this? Many people after taking the first dosage, do not end up taking the second dosage. So you too, should not take second dosage!’
Father Savvas: “She gave him some courage.”
[Priest-Monk]: “As she was telling me these words, I felt a certain refreshing dew entering my soul.”
Father Savvas: “This was from God.”
[Priest-Monk]: “I was consoled by God’s grace.”
Father Savvas: “Because he had begun to repent. God sends people to console us, to inform us. This is how God works during such situations. Just a word, though many times irrevlevant, shows us the way.
So she said, “Ok, you made a mistake. Don’t make the next mistake.” i.e. don’t take the second dosage.” Let’s continue:
[Priest-Monk]: “I find it unnecessary to mention the despair I went through and the tears I shed. I don’t know whether it’s a coincidence or not, but exactly 40 days after the vaccine I started to feel the grace of God again.”
Father Savvas: “After 40 days he began to feel that he was baptized again. He came back with repentance.
So he confessed his sin, received the rite of forgiveness, and shed many tears. As it is written here; He cried continuously for 40 days and only then did he begin to feel the grace of the Holy Spirit.”
[Priest-Monk]: “I began to feel peace and consolation, that God had forgiven me for what I had done, even though I had no knowledge of what this vaccine actually was.”
Father Savvas: “Sadly there are many people like this, not everyone is indifferent, there are others who are directed by tyranny and fear… or by pressure, by their children or doctors, etc.
However, when you have the correct information you don’t submit to all of this. This poor fellow was seduced, but now he has corrected his actions.
So after 40 days he began to feel the forgiveness of God.”
[Priest-Monk]: “I do not dare or want to know what would have happened to me if I had taken the second dosage of the vaccine. The only thing I can say is that God felt sorry for me.
Even though now I feel better, I have not recovered to the state I was in prior to taking the vaccine.
In my humble opinion, this vaccine by Pfizer that I took is a mark (seal), but not the final mark. Most likely a forerunner for the final mark of the beast (Book of Revelations).”
Father Savvas: “This last statement correlates with another remark made by a demon during another exorcism. A close friend of mine, a respectable Priest-monk had told me. He had a dialogue with a demon during an exorcism.
The demon told him, “Yes, this vaccine is not the final mark (stamp), but it is still a mark, a forerunner… And those who are like you, when they take the vaccine they lose their light.” The demon continued, “A short man had burned us with these exorcisms!” This short priest (Elder) is well known and performs exorcisms… The demon continued, “Now that he (the short Priest) took the vaccine, he has lost his strength. Now I am able to approach him and kiss his hand!”
Previously, the demon would tremble with this Elder. But now that the Elder has taken the vaccine, the demon is able to get his blessing. This correlates with the Priest-monk we have been reading about: “Even though now I feel better, I have not yet recovered to the state I was in prior to taking the vaccine.”
As you can see, these vaccines not only cause physical (biological) harm, but spiritual harm also.
We have read this testimony as an example, and it is from a person living a proper ecclesiastical (church-minded) life. Because as we previously said, when someone makes a mistake that is impacting many others, it’s most beneficial to correct this mistake publicly. This is in order to protect other members of our Holy church from making the same mistake.
As St. Chystostomos says, “The lukewarm Christians are living in comfort.” The lukewarm are those who want to combine everything; the world, Christ, hedonism (love of sexual pleasure), avarice (love of money), the external appearances (vanity)… to not be disenfranchised (segregated), go to Church, take Holy Communion, Holy Confession, etc. These lukewarm “Christians” cause the most damage to the church.
And they don’t admit to their mistakes, as they think they do everything correctly. If they make a mistake, they don’t correct it. They do not publicly repent of their sin so they may protect those around them.
As it is written in the book of Revelation, these are people that God will spit out (vomit them out). It is best to be hot or cold, never lukewarm. The one who is spiritually cold may at one point understand their spiritual blindness and become hot. God wants us to be hot. However, the lukewarm are comfortable.
Sadly, most people nowadays are lukewarm. As mention by Father Athanasios Mitilinaios, most Christians are lukewarm. We too are lukewarm and need to stop being lukewarm.
To a faithful person of God death does not exist, this is the reality. We have forgotten this and we presently fear death. Not only do we fear death, we also fear being fined, possible imprisonment, and prosecution. In NO case can a person call themselves a Christian if they fear death. When a person fears dying, they become an idolater or an atheist.
Instead a Christian should long to die. The saints wanted to die. The reason why Christians truly want to pass away is so they can be fully united to the Lord they worship and love above all else. They want to go and are joyful when they are passing away. However, they never cause death to themselves, they do not commit suicide. But when the opportunity arises to become a martyr and confessor for Christ, they do this without fear of death.
Unfortunately, these things are not being heard from the Preachers, Bishops and Priests, but as you know are heard from the demons.
A well-respected Priest-monk who perform exorcisms, once told me what a demon said to him. The demon said to him, “How are you Christians fearful of death? I have seduced and deceived you into taking the vaccine with the fear of death!” A demon again confesses that the vaccines are his doing. In fact, he says, “We did a ceremony at a Lodge in America for the vaccines.”
See what the demon confesses! The demon continues: “What did you fear? For you (Christians), death does not exist.” For Christians, death does not exist. Do you understand, fellow brothers and sisters, where we’ve come to?
In the Gospels, Christ said something correlating with our present situation. When He entered Jerusalem and the children were calling out, “Hosana! Blessed is He who comes in the name of the Lord!” The children were proclaiming Jesus as the awaited Messiah. And others who were indignant were saying to Jesus, “Are you hearing what they are saying to you?” Christ responds, “I tell you that if these should hold their peace, the stones would immediately cry out.”
So correlating with today’s situation, now that the Priest and Archpriest are not proclaiming that death has been conquered, the demons are proclaiming it instead: “How are you Christians fearful of death? And are all running to take this vaccine?”
Which isn’t really a vaccine, but rather a gene therapy/technology, with the aim of oppressing humans. This is the aim, which is why there will be more dosages. In fact, there are many people who say there will be 7 dosages. The demons also state that there will be 7 dosages.
Those who were deceived into taking the first or second dosage, hopefully, will not take the remaining dosages. May they repent, weep and go to confession, because their salvation is in danger. We all ought to be vigilant of this.
I’ll read another Priest-monk’s testimony to you all, which was published on the internet unless it’s now been erased. Because whatever is true these days, is usually censored on the internet. As you know the internet is Satan’s. Unfortunately, the internet is not governed by God’s people. Whatever they want they remove, as much as possible they eliminate anything that is good and right.
So this Priest-monk who published his testimony on the internet said, “I considered taking the vaccine was nothing. So I went and got it. But after I got it, I lost my prayer. I felt Satan coming and hugging me. For 40 days I could not even say ‘Lord have mercy.’ ” This Priest-monk has testified this publicly. I personally know him. He is from a Monastery in Peloponese. He also urges everyone not to make his mistake (of getting vaccinated), and whoever has already taken it, to not take any further dosages, which will cause more harm for both body and soul.
In conclusion, these are the things I wanted to say to you, brothers and sisters in Christ. As you can see, there are many testimonies from many individuals. God even caused the demons to give a testimony. Remember what the demon was saying to Priest-monk during the exorcism?
“Why am I telling you all this?! I do not want to tell you, but I am being pressed.”
We must also remember that the devil lies too, however, there are many times when the devil says the truth, especially when forced with the prayers that are read during an exorcism. Likewise, we read in the gospels, the demons saying the truth to Jesus, “You are the Son of God!” The demons confessed that Christ is God.
Let us take these things into account so that we don’t fall into this trap of Satan. Which as you can see, Satan is using much force to direct all of humanity into this trap. So he can kill as many as possible. This is the murderer (man-killer) that Satan is. This is now all coming into fruition with what is happening globally.
The devil wants to take as many souls as possible, this is his final goal. If all these things (vaccines) were good why would they make them mandatory? Something that is good is not forced. You see Christ, whenever He went to heal someone, He would ask, “Do you want to become well?”
These rulers nowadays are trying to force us to be well. That’s what they think. However, they don’t want to make us healthy, quite the opposite is occurring actually. Because we know these vaccines cause sterility and thousands of other adverse reactions. Over 2 million adverse reactions were recorded in Europe. And over 600,000 adverse reactions in America. Over 21,000 deaths in Europe, and 14,000 deaths in America. These are all mainstream statistics that you can find online.
Therefore we do not need to eat idiotic grass and sleep with our boots on (greek expression; We don’t need to be idiots). Under no circumstances are we to accept these things that the evil one is selling us.
May God bless us all. May John the Baptist guide us. To not only cease committing evil, but to call out evil when we see it. Whoever cowers and is silent before sin, becomes an accessory and an accomplice to sin. So what I tell you, you need to tell others, inform all our siblings so that they may not fall into this trap by Satan, and lost the kingdom of God. Amen.”
Tuesday, September 14, 2021 (Exaltation of the Holy Cross)
Jesus said: “My son, you are fortunate to have been given a relic of My true cross that you wear so you can pray over the people. The first reading of Moses holding up the bronze serpent on a pole, is a precursor for when I would be held up on the cross to redeem all of humanity who accepted Me. This was a nice service that your priest did to honor My Holy Cross. I am an example to follow that you each take up your own cross in life to carry out the mission I have given to each of you. In life you all are struggling with the trials of life with your health and working to make a living. Call on Me to help you through the hard times of deaths and sickness. Even now your authorities are trying to force a poisonous Covid shot on everyone. Stand up to these evil people, and refuse these shots that could kill you. This is illegal for the evil ones to force a death shot on people to reduce the population. Trust in Me to protect My believers from these unnecessary death shots. Eventually, you will need to come to the protection of My refuges, when these evil ones will try to kill you.”
Monday, September 13, 2021 (St. John Chrysostom)
Jesus said: “My people, you repeat the Centurion’s words at every Mass when you say: ‘Lord, I am not worthy that you should come under My roof, but only say the word and my soul shall be healed.’ I was so pleased with this man’s faith that he should be an example for all of you to follow in humble worship of Me. Many of you may have wished to be with Me when I was on the earth, but you do have Me in My Real Presence in the consecrated Host. You will see Me soon in My coming Warning, so have your soul prepared with frequent Confession. The unvaccinated people will be further harassed by Biden in his evil effort to get everyone vaccinated with the poisonous Covid shots. You are hearing of young adults dying without explanation. Now, your morticians are seeing dead bodies with many blood clots all over their body which is a result of the Covid shots. This is not being made public, but some people are speaking out of such deaths. You will be protesting the Covid shot mandates for the nurses at your local hospital today. There has not been much resistance to Biden’s vaccine mandate, and this is one instance. Continue pushing back against these mandates, and advise people to not take these poisonous Covid shots that ruin your immune system, and could kill people in a few years. Trust in Me to protect My believers when the authorities will try to kill you. Then I will call you to My refuges of protection.”
Jesus said: “My people, after I have cleansed the earth of all the demons and evil people, I will truly renew the face of the earth, and My faithful will see a new heavens and a new earth. I will renew the face of the earth as My faithful will be raised up into the air. Once I have renewed the earth, I will call My faithful down into My Era of Peace. You will live a long time and you will be young again. You will have no evil present, and you will grow in holiness. Once you die, you will be as a saint, and you will be brought into heaven. This will be your reward for being faithful to Me during the tribulation. Trust in Me so you can face your trials of the coming tribulation.”
Sunday, September 12, 2021
Jesus said: “My people, you remember how St. Peter answered: ‘Thou are the Christ, the Son of the living God.’ It was the Holy Spirit that inspired St. Peter to know and respond in this way. My people also have the Holy Spirit making My Real Presence known to you, and you have accepted in faith that I am Present in the consecrated Host. You also accept on faith how I died to redeem sinners, and I was resurrected on the third day to show you how My faithful will also have their souls rejoined with their glorified bodies on the last day. In the second reading you read how you cannot have faith without works. If you truly love Me, then you must love your neighbor and show it in your actions of good deeds to help them with food, donations, and sharing your faith. By trusting in Me for everything, I will lead you on the narrow road to heaven.”
Saturday, September 11, 2021
Jesus said:“My people, the one world people were instrumental in taking down the two trade towers and Building 7 that was not hit by the planes. This was a traitorous act that killed so many innocent people, and the dust harmed even more people. You pray for their souls and the families who mourned the loss of their loved ones. These are the same evil ones who are trying to reduce the population with their man-made viruses and toxic Covid shots that they are using to kill people. You know they are evil by their evil actions, and they are like the rotten tree that bears rotten fruit. My good people, who follow Me and avoid these Covid shots, are the good trees that bear good fruit in their actions of worshiping Me and doing good deeds for their neighbor. Trust in Me to protect you from the evil ones at My refuges, when they will try to kill you.”
Jesus said: “My people, you have seen some powerful storms like Hurricane Ida, where the damage is still being assessed. You are also seeing more fires out West taking their toll on your homes. The second vision of icicles on a roof is a sign of a colder than usual winter that could cause more damage. Some of these weather events are a punishment for your sins of abortion and your sexual sins. You could see a possible increase in your unemployed workers because they will refuse to take the Covid shots and they know how life threatening that the poisonous shots are. Biden is going along with the one world people’s plan to reduce the population with the mandatory Covid shots. Events will soon try and force the mark of the beast on your people, but do not take the mark of the beast, or you will lose your souls to the evil one. Trust in Me to bring My Warning before vaccinated people will die from the next deadly virus. After the six weeks of conversion, I will call My people to My refuges.”
Friday, September 10, 2021 (Eastridge Class ‘61 deceased intention)
Jesus said: “My people of the Eastridge Class of ‘61, I am happy you are all here to see each other again. You have seen the list of your deceased classmates, and you are sorry they have passed on. You are seeing in the vision how some souls of your classmates are in purgatory, and they are grateful for the graces that they received from this Mass intention. Remember to keep praying for them, as their souls can move closer to heaven with your prayers and Masses. Many people think souls go directly to heaven, but most souls need some reparation for their sins in purgatory. These souls are thankful for remembering them.”
“My son, you are seeing many people struggling with your recent Biden Covid vaccine mandate on workers. Not everyone realizes how these shots can ruin your immune system, and can even kill you, especially when a more deadly virus is released. These viruses and vaccines were purposely made to reduce the population by the one world people. So refuse to take any Covid shots or flu shots, even if you may lose your job. Biden is treating you as if he is a dictator, and he is forcing his control over everyone. Be prepared to come to My refuges when your lives are in danger.”
Jesus said: “My people, initially you saw how the government was trying to force your employers to have their employees take a Covid shot, or they could be fired. Employers were a little apprehensive to push this forward. Now, Biden is back to using Executive Orders to try and get the unvaccinated workers forced to take the Covid shot. There will be a challenge to this order because there is not a lot of recent deaths to justify such an overall order. The previous lock down did not slow down the spread of the virus, and now being vaccinated does not appear to stop the delta variant either. There is no true science behind this order, and it is only control over the people that Biden is pushing. These shots will endanger your people’s health because these viruses and vaccines are meant to kill people and reduce the population. Refuse to take the Covid shots because they will ruin your immune system, and vaccinated people could die from the next deadly virus. You are having a hard time finding workers, but if you fire those people, who refuse the shot, you will have even more unemployment and businesses could fail or go bankrupt. Vaccine mandates could ruin your economy, so they should be rejected. Pray for your people to find a way around these mandates.”
Thursday, September 9, 2021 (St. Peter Claver)
Jesus said: “My people, today’s Gospel speaks about loving your enemies and sharing what you have with them. It is hard to forgive someone who has cheated you out of money, or who has harmed you unjustly. You love to share what you have with those people in need, but you are hesitant to help those people who do evil things. I shine My Light and graces on all people, but it takes more grace to love your enemies. You can at least pray for your enemy to help save their soul. You do not want to see anyone go to hell, so this is the least you could do to help your enemy. Every soul is lovable, even the worst sinner, because they are made in My image. I want My people to reach out to all souls in your evangelization efforts, and I will repay you. When you come to your judgment, I will ask you if you loved Me and all of your neighbors, even your persecutors. By loving everyone, you will gain your salvation.”
Jesus said: “My people, you are seeing the beginning of the vaccine mandates that will be getting so bad, that I will soon have to bring My Warning and the refuges to save your lives. Today, it is mandated shots for workers not long after Labor Day. Tomorrow, they will try and come door to door to demand the same shot. If they can mandate Covid shots and boosters to everyone, then they will next demand that you take the mark of the beast, and you will be faced with losing your soul. So absolutely, never take the mark of the beast no matter how they threaten you, or you will lose your soul. This will be followed by a persecution of those people who refuse to take the mark of the beast, meaning they will try to kill resisters in the detention death camps. Before they try to kill My faithful, I will bring My Warning, and later I will call My believers to My refuges of protection. This is why I have been alerting My refuge builders to be ready to receive the people that I will send to them.”
Jesus said: “My people, you have heard from your friend how evil the mandates are in Puerto Rico that the unvaccinated people cannot buy groceries or gasoline. This Hitler style shutdown is coming to the rest of America, so if your people do not fight these mandates, then you will lose all of your freedoms. When your lives are in danger, I will call you to My refuges.”
Jesus said: “My people, earlier in the year you had a shutdown of your churches, when you could not come to Mass nor Holy Communion. You had to use your internet to see a Mass, but you could not have My Eucharist. These mandates are following the atheistic persecution by your socialist communist government who hate Me, and they will restrict your worship of Me. Eventually, even the internet will shut down religious services. Trust in Me to lead you to My refuges when your lives are threatened.”
Jesus said: “My people, I have warned you about the possibility of the evil ones bringing a worse Covid virus that could cause vaccinated people to die quickly. This is the vision that I gave you of the dead bodies lying all over. These Covid shots are ruining your immune system, so when this deadly new virus comes, people will be dying all over. This has been the plan of Satan to reduce the population using viruses and vaccines. I am giving you My Good Friday oil and the exorcism water to drink for the vaccinated people so they will not die from the vaccine. As you are threatened with this new virus, I will bring My Warning first and then My angels will lead you to My refuges.”
Jesus said: “My people, I have promised you that I will bring My Warning before the next deadly virus will be released. On the day of the Warning you will see two suns in the sky, and there will be some days of darkness. You will face your life review, a mini-judgment, and a taste of your present destination. After the Warning, you will have six weeks of conversion. You will be warned not to take the mark of the beast or the Covid shots. You will be returned to your bodies, and My believers will be called to the safety of My refuges.”
Jesus said:“My people, I have warned you in previous messages that I would give everyone an inner locution when it is time to come to My refuges. After you receive this inner locution, I want My faithful to pack up your backpack, tent, and sleeping bag, and leave your homes for My refuges within twenty minutes. Call on Me and I will have your guardian angel lead you with a flame to the nearest refuge. As you leave your home, your angel will put an invisible shield over you so the evil ones cannot see you. You can enter My refuges if you have a cross on your forehead. My angels are marking a cross on the foreheads of all of My believers. The refuge angels will not allow anyone without a cross to enter. When you look on the luminous cross in the sky at My refuge, you will be healed of all of your illnesses. Give praise and thanks that I am protecting your lives.”
Jesus said:“My people, I will lead My faithful to My refuges, but they will enter slowly to give My refuge builders time to assign beds and places for each believer. Be patient with My refuge leaders because they will need to know your skills so you can be assigned jobs to help all of the people at each refuge. Leaders for various jobs will be chosen, and each leader will gather helpers to carry out their mission. You will have two meals every day, and each person will be assigned Adoration hours for the day. My believers will need to be patient because it will take time to organize your jobs. I will multiply your food, water, and fuels. Fear not because you will have powerful angels side by side all around the perimeter of your refuge to protect you. You will have daily Mass by a priest, or My angels will distribute daily Holy Communion. Your daily prayers, Holy Communion, and Adoration hours will keep you close to Me throughout the tribulation of less than 3½ years.”
Wednesday, September 8, 2021 (Nativity of the Blessed Mother)
Our Blessed Mother said: “My dear children, I bring the rain as at Fatima, Portugal, as the Lord’s rain is needed in your land. It is also symbolic of how my Son, Jesus, rains down His graces on all of His children. You are celebrating my birth, but the readings speak of my heritage from King David, and the birth of my Son, Jesus. He is a Son of King David through my ancestry. There is also a mention of my Son’s birth as the dawning of salvation, because truly His death and Resurrection have brought salvation to all of mankind who accept Jesus. I am the instrument to bring Jesus into this world, and because of my place as the Mother of God, the Lord has called me blessed. My Son and I have our two hearts joined together, and I bring you to Jesus as a spiritual mother as well.”
Jesus said: “My people, your media is constantly attacking the unvaccinated, and your government is doing everything to get your employers to force workers to get vaccinated. You should not be forced against your will to take a poison Covid shot, and even the vaccinated should not take any booster shots. You are entering the flu season, and it is hard to see your tests telling the flu shot apart from the Covid virus. You need to keep your immune systems protected with vitamin C and D3 2000. Take some zinc as well as your Hawthorn pills. By eating properly with occasional fasting, you can stop any Covid delta variant attacks. Refuse to take any Covid shots or flu shots. The restrictions on the unvaccinated will be getting worse, and the evil ones will use any increases in Covid cases for another potential lock down. I want My refuge builders to be prepared for taking many people into their refuges. The time to call people to My refuges draws close, as your lives will be in danger soon from any shut downs or more mandatory restrictions. Have your food and water ready to share with the people. Trust in Me and My angels who will be multiplying food and fuels for your people to survive.”
I, “a soul”, was intending to make a 2-part video series for MaryRefugeOfSouls, but this subject matter is SO IMPORTANT for everyone to receive today, I have decided to do a blog post with the vital information first. Then, I will make the 2-part video series later when I have time.
What is the subject matter of this post?
It is about Graphene Oxide, a toxic poison made of lead found in all the Covid-19 vaccines. It is a unique electromagnetic material that can also interface with the human brain, which will form the basis for the mind control properties of the mark of the beast mentioned in Sacred Scripture (Book of Revelation).
This blog post consists of both parts that I intend to use for the future video series. Each part has first quotes from messages given to Catholic prophet, John Leary, from Our Lord, Jesus Christ (http://johnleary.com). Then, two brief videos follow each prophecy segment illustrating the scientific evidence related to the quotes from Jesus.
My hope is that blog followers of MaryRefugeOfSouls will review this entire post, including watching all the videos. Then, they will, in turn, share this important post with all their loved ones — sharing this post far and wide so everyone can have this critical information that is a life-and-death matter for all concerned today.
Thank you for caring. God bless you all and all of your loved ones.
I love you, –a soul
SHOCKING GRAPHENE OXIDE
PART I – GOD REVEALS THE TRUTH – ABOUT THE DEADLY POISONS IN ALL COVID-19 VACCINES (includes scientific evidence)
PROPHECIES, PART I
Our Lord, JESUS CHRIST, Speaks About Graphene Oxide Poison Found in ALL Covid-19 Vaccines
Prophet John Leary Friday, July 30, 2021
JESUS CHRIST said:
“My people, it has been shown by some doctors that ALL THREE COVID SHOTS ARE POISONS that are not necessary to take because 98% of the people do not die from the Covid-19 virus.
These shots contain GRAPHENE OXIDE and iron oxide additives along with chemicals that can permanently cause your body to manufacture the SPIKE PROTEIN that can eventually KILL people…
You will soon see the vaccinated people spreading the virus, and they will be getting sick as time goes on…
Trust in My protection from these POISONOUS SHOTS that will RUIN YOUR IMMUNE SYSTEM if you take them, and they could eventually KILL you.”
Thursday, August 5, 2021
JESUS CHRIST said:
“My people, I have told you that the DEMONS GAVE THE KNOWLEDGE to make the Covid virus and the Covid vaccines to your scientists.
So this is why you are seeing the DEVIL SPREAD DEATH all over the world using the Covid vaccines. (Reference Note by a soul, Pale Green Horseman of the Apocalypse)
These shots have POISONOUS GRAPHENE OXIDE, iron oxide, and lipids that produce the SPIKE PROTEIN all over your body.
Gradually the constant inflammation produced by these shots WILL KILL the VACCINATED PEOPLE in a few years, unless they are healed (by God).”
* * *
The following 2-minute video from StewPeters.tv shows expert, Dr. Jane Ruby, reveal scientific proof that the Covid-19 vaccines are 99.9% poisonous (containing 99.9% Graphene Oxide).
VIDEO 1 GRAPHENE OXIDE – Dr Jane Ruby – 99 Percent POISON IN VACCINE (2:09 minutes)
The following 3-minute video from StewPeters.tv shows expert, Dr. Jane Ruby, reveal REAL microscopic evidence of permanent physical damage to human blood due to graphene oxide poison.
VIDEO 2 GRAPHENE OXIDE – Dr Jane Ruby – Blood Smear VACCINATED BLOOD DAMAGE (3:12 minutes)
* * *
SHOCKING GRAPHENE OXIDE
PART II – GOD REVEALS THE TRUTH – ALL COVID-19 VACCINES ARE ONE STEP FROM THE MARK OF THE BEAST (includes scientific evidence)
PROPHECIES, PART II
Our Lord, JESUS CHRIST, Reveals ALL Covid-19 Vaccines Are Precursors to the Mark of the Beast
Prophet John Leary Saturday, October 3, 2020
JESUS CHRIST said:
“…In fact this vaccine will be worse than the disease, and it will be combined with a chip in the body to tell if you had the vaccine.
There is even a threat that the evil ones may use nano particles to cause a control of your body.
This chip or nano particles will be the mark of the beast.
So do not take any vaccine and do not take any chip in the body, because it will control your mind.
Refuse to take any flu shot. Refuse to worship the Antichrist, and avoid taking a virus test if possible.
The authorities will try and kill all of those people who do not take the vaccine and the chip in the body.
Also refuse to use 5G devices because the microwaves will harm your body…”
Monday, December 21, 2020
JESUS CHRIST said:
“…I have told you many times DO NOT TAKE THIS VACCINE, EVEN IF THE AUTHORITIES THREATEN TO KILL YOU.
The NANO PARTICLES will CONNECT you by 5G MICROWAVES to the CLOUD that will transmit your bio information to those people who want to control you.
These vaccines will PERMANENTLY CHANGE your DNA as the NANO PARTICLES are like LITTLE ROBOTS going through your body.
You are risking DEATH or ALLERGIC REACTIONS with this vaccine, SO DO NOT TAKE IT.”
* * *
The following is a 5-minute video featuring a pharmaceutical expert, Karen Kingston, speak about Graphene Oxide being a trade-secret poison found in all the Covid-19 vaccines. She also speaks about Graphene Oxide having neural (brain) interface properties that can be used for mind control in people.
VIDEO 3 GRAPHENE OXIDE – Former Pfizer employee – Karen Kingston POISON – MARK OF THE BEAST (4:53 minutes)
The following is an over 3-minute video that demonstrates how Graphene Oxide particles work actively together to interconnect like little robots. Now imagine this evil-conceived matter being in your blood traveling throughout your body and organ systems.
VIDEO 4 GRAPHENE OXIDE – Graphene Oxide Alive ELECTROMAGNETIC – MARK OF THE BEAST (3:24 minutes)
* * *
CONCLUSION
The following is a brief video with some summarizing thoughts to consider from independent media journalist, Stew Peters.
CONCLUSION – Would you put this in your body? (1:30 minutes)
Jesus said: “My son, you have been out evangelizing people so they could be converted for over 25 years. Now the time to bring people to Me is growing short before the coming Warning and the tribulation of the Antichrist. Your mission has been to help save as many souls as you can, and the time to save people from hell is running out. The devil and the media are controlling the minds of many people, but My power is greater. So call on Me and the Holy Spirit to help you every day to have people understand that they are both body and soul, and made to My image with a free will to choose between heaven or hell. My Warning will wake up every person to realize how I will judge each life with your life review and your mini-judgment when people will have a real taste of their future destination. If you love Me and repent of your sins, you will be on the narrow road to heaven. But if you ignore Me and refuse to repent of your sins, you will be on the broad road to hell. In heaven you will enjoy being with the One you love forever. I have shown you hell where the demons are tormenting souls and the souls there are like black embers who are suffering the flames forever with no hope of escaping. Those people, who are on the road to hell, will have a taste of this suffering, so they can see what their life is choosing. My faithful will have a chance during the six weeks of conversion to try and convert these souls when there will be no influence of the devil. Pray for the conversion of souls and reach out to give them My Blessed Mother’s brown scapular, so she can keep them from suffering the flames of hell when they wear and believe in the brown scapular’s promises.”
Monday, September 6, 2021 (Labor Day)
Jesus said: “My people, America needs to wake up out of your media brainwashing, and see how you are a bunch of frogs being slowly boiled to death in water. Your jobs are being threatened by a vaccine mandate that will kill you, and you should not take it. If you are dead, what good is your job? When are your people going to rise up in a major protest, or strike against your employers? If you do not fight this vaccine mandate and passports, you will soon be like Puerto Rico where unvaccinated people cannot work, buy groceries, or gasoline(* see endnote). You are in the middle of a communist takeover, and you do not even realize it. If these evil ones can force you to take an unneeded, poisonous, Covid shot, then they will force you to take the mark of the beast next. You cannot take the mark of the beast, or you will lose your soul. So it is better for America to resist these mandates now, or you will lose all of your freedoms. Once you cannot buy food or gas, and they threaten your lives, I will bring My Warning, and I will call you to My refuges. If your people do not resist, then I will bring My Warning, and your free country will cease to exist. Fight for your freedoms now, or you will lose all of them. Trust in Me to protect My faithful at My refuges of protection with My angels. If you quit your job, you can use your three months of food to survive a while.”
Jesus said: “My son, you are planning your Zoom interview for TV tonight that you will be giving tomorrow. Call on Me and the Holy Spirit to give you the words to speak to the people. It is good to get My message out to the people who may be influenced to be converted. The people need to know that it is important to choose your destination where you will be after you die. They will either follow Me to heaven, or follow the devil and the world to hell. I do not want to lose even one soul, so make it clear that this choice will be forever, and not just a lifetime. You also will be traveling again to support a young man and his mother, as the exorcist priest will be doing another exorcism session with him. You can do some fasting and prayers in support of the priest. Pray the long form of St. Michael’s prayer going and leaving from this exorcism. You continue to bring your relics for additional support. I have asked you to continue to support this family as often as you can. Trust in Me to guide you and protect you from any demonic influence.”
Sunday, September 5, 2021
Jesus said: “My people, there are many places where you feel out of your comfort zone with different groups of people. But I reached out to foreigners, lepers, fornicators, and even tax collectors. When you are sharing My words and trying to convert people to a Christian way of life, you need to mix with people where you do not normally visit. When you share My love and reach out to help people, you have to make time for them, even at your own inconvenience. You can see how the evil leaders are trying to create divisions in your country over whether or not you are in favor of abortion, euthanasia, drugs, and now over being forced to take an unneeded Covid shot. Most of these divisions center around the means of the death culture that has little regard for life. When you love people and want to offer help, you should not be oppressed by evil people who want to force their will on you and control your lives. Because of this forced control, and desire by some to reduce the population, I will soon be calling My faithful to the safety of My refuges, as your lives are being threatened. Trust in Me to protect you from harm as My angels will defend you. Call on My help when you are in need, or when you are being threatened.”
Saturday, September 4, 2021
Jesus said: “My son, I thank you for all of your driving and giving your talks last weekend. This accident you had with your knee, was an attack by the demons who did not want you to travel to these talks. Sometimes you have to suffer some pain for the souls you are helping with healings and spiritual help for the people. Keep struggling in your efforts to help save the babies at Planned Parenthood today, and protesting the vaccine mandates at the hospital. Abortion and the vaccine mandates are the two greatest intentions you should be praying to stop. Even if you have to suffer pain and inconvenience, these causes are well worth your evangelization efforts. You may suffer this struggle to save the babies and fight the forced Covid vaccinations, but it is all worth your time and dedication. Trust in Me that you will not have to wait long for My justice to fall on the evil ones behind these injustices.”
Jesus said: “My people, trying to force employers to have mandates to have a Covid vaccine for their employees or they would lose their jobs, is against your country’s freedoms. Everyone should not be forced to take these poisonous vaccines against their will. The threat of losing your job should not be conditioned on taking these toxic shots. If you have a choice between dying with these shots or making money from a job, your best choice would be to quit that job. If all jobs are facing this mandate, then you need to rebel against this. If your government can force you to take a shot that will kill you, they can force more boosters and the mark of the beast on you as well. It is better to refuse these death shots and for sure to refuse to take the mark of the beast because you are risking the loss of your soul. People need to know that the true doctors are saying that these poisonous shots could kill you in a few years, especially if the vaccinated people come in contact with another wild corona virus. At one point these evil ones want total control over your lives, and the Antichrist will control all of the jobs and the networks, so it is better to quit now. This is not an easy decision, but once they impose a total lock down on the unvaccinated people, as in Puerto Rico, you will have to fight these edicts or die of starvation. Unvaccinated people in Puerto Rico cannot even buy groceries or gasoline(* see endnote). If you are locked down, pray to Me and I will multiply your food. This intolerable lock down will precipitate My Warning, and I will call My people to My refuges. Be prepared soon to leave for My refuges because the evil ones will soon try to kill the unvaccinated people because of your resistance. Trust in Me to help vaccinated people get healed with the Good Friday oil or the exorcism water. They must believe in My healing power to heal them. My justice will fall soon on these evil ones for planning to kill many people with their vaccines.”
Friday, September 3, 2021 (St. Gregory the Great)
Jesus said: “My son, this first vision of an all encompassing black abyss represents all of the evil people who are perpetrating the viruses in the chemtrails, and the more sinister poisonous vaccines that they are trying to force on everyone. Your evil health leaders are having employers threaten people to lose their jobs, if they do not take these useless vaccines that will kill people in time. Some employees may be able to take a weekly Covid test and avoid the poisonous vaccines. These evil health officials are passing edicts that are nothing but lies. There are as many vaccinated people having sickness and complications, as the unvaccinated people are suffering from flu and Covid sickness. This darkness of control will lead to mandates for the mark of the beast. Absolutely refuse to take the mark of the beast, even if they take away your job or threaten your life. There is no cure for the mark of the beast and those people, who take it and worship the beast, will be lost forever in hell. When these evil ones mandate the mark of the beast, you will see My Light to lead you to My refuges of protection. You will endure the tribulation of the Antichrist for less than 3½ years. At the end of this reign of the Antichrist, I will destroy all of the evil people and demons, and they will be cast into hell. My believers I will bring into My Era of Peace. Do not fear this evil because My power is greater, and these evil people and the demons will be cleansed from the earth into hell. Stay true to following Me, and you will be rewarded in My Era of Peace.”
Jesus said: “My people, time is running out for Satan and the demons. You are seeing a pivotal battle between My angels and the demons. At the end of the tribulation, I will come to help My angels defeat the evil ones at the Battle of Armageddon. I told you that I and My angels are more powerful than Satan and the demons. This will be a crucial battle that forces of good will defeat the evil forces. After My victory, all of the evil people and the demons will be cast into hell. I will then renew the earth and I will bring My faithful into My Era of Peace. Trust in Me that Satan and the demons will face a crushing defeat. Trust in Me to protect My believers at My refuges.”
Thursday, September 2, 2021
Jesus said: “My people, at times you will come up against situations that seem impossible to overcome by your own ability and strength. It is at that time that you need to call on Me to help you, because I can do the impossible. St. Peter had fished all night without success, so when I asked him to lower the nets for a catch of fish, he was reluctant. But to please Me he lowered the nets. They brought in so big a catch of fish that they needed help with another boat, and the boats were both so filled, that they were almost sinking. St. Peter was so amazed at such a catch of fish, that he felt that he insulted Me for being reluctant to lower the nets again. This is when he said: ‘Leave me Lord, for I am a sinful man.’ You have an almost impossible situation now, when your leaders are forcing the employers and colleges to mandate Covid vaccines for all of their employees and students. This is against the science of health when these poisonous Covid shots are toxic to your body and they will ruin your immune system. You have tried to fight these mandates with exemptions, but your New York governor is not allowing religious exemptions. You need to give this over to Me, and I will lead you to My refuges of protection when your lives are in danger. People do not want to lose their jobs, but they do not want to take the poisonous Covid shots either. You are seeing devastating weather damage from Hurricane Ida that could stop jobs and shots in some areas. Trust in Me to find a way to defeat these evil mandates that are lies and are not needed. In some cases you can have Covid virus tests each week, but this is just a harassment for the unvaccinated people.”
Jesus said: “My people, I mentioned before how Hurricane Ida was a punishment for your abortions and all the sins of your people. In this vision I reminded you of how people came to church on 9-11-01 right after the plane crashes to repent of their sins. You have just had some terrible destruction in Louisiana and other states from rain and tornadoes. You still are uncovering people who died from this storm. You all could come to church to repent of your sins and come to Confession. You could still have more disasters as your fires and drought out West. If your people repent, I could hold back My justice, but if you do not repent, then you will see more destruction.”
Jesus said: “My people, just as you were asked to help Haiti, now you will be asked to help your own people in Louisiana to provide electricity, gasoline, water, food, and shelters. Even in your own state of New York and neighboring states, you are recovering from record flooding. This storm has touched many people with some destruction. As this storm came on the same day of the year as Katrina, this is a sign for your people to repent and improve your spiritual lives.”
Jesus said: “My people, on your TV news you are seeing pictures of the devastation all over the Southern states. You have had declarations of emergency so Federal aid can be brought to the areas that need help the most. People, who lost their homes, will have to start over and hope that insurance and government aid can provide a new home. These people are needing fresh water and food. Pray for these people to find what is needed for their survival.”
Jesus said: “My people, many Americans and Afghans were fortunate to be airlifted to safe places from the Taliban. You may be asked to help refugees from Afghanistan in addition to all of the illegal immigrants who are being sent to cities all over your country. The open Southern border is causing many problems with sick people and finding shelter for these people. Pray for your country that is suffering from storms and fires.”
Jesus said: “My people, your employers and colleges are trying to enforce vaccine mandates for their workers and students. It is hard to legally enforce Covid vaccine mandates when Covid shots are still considered experimental. There is some misunderstanding if the FDA can approve mandating shots that are toxic to your bodies. Any such approval has not gone through the necessary testing. Refuse to take these poisonous shots and boosters that have not been proven to be safe and effective.”
Jesus said: “My people, there is talk that the teachers may require Covid shots or weekly testing for the Covid virus. The younger children have not been getting sick, but it is hard to know if the children will need Covid shots. Many parents do not want their children vaccinated with the poisonous Covid shots. You are seeing more home schooled children to avoid having to take a Covid shot. Pray that your children and grandchildren are not forced to take the Covid shot.”
Jesus said: “My people, your people need to be understanding of the fuel shortages until your places of production can come back online. Your country should control the prices because of a short supply of some fuels so people are not gouged with high prices. Your people might have to share fuels and staples so your stores do not run into low stock on your store shelves. Hoarding and price gouging should be controlled until supplies are normal again. You will be seeing more food shortages as time goes on, so be prepared with your three month supplies of food that you were supposed to have. You may even have to call on Me to multiply your food where you have continuing shortages, especially of fresh water. Pray that your families can have what they need for their survival.”
Wednesday, September 1, 2021
Jesus said: “My people, since Adam’s sin, you are all appointed to die one day as a consequence. You also have a weak body to disease and viruses, and you can get sick as well. You have the evil ones spreading flu like diseases in the chemtrails that are being tested as Covid virus, when any corona virus will give a positive test. It is one thing to be quarantined for 10-14 days if you have a flu or Covid virus, but making people take a poisonous Covid shot when you already have antibodies, is nothing but communist control like a dictatorship. Your health people do not even allow sick people to receive ivermectin or hydroxychloroquin because it is cheap and the one world people want to reduce the world population. To counter the death culture people, I have given you a blessing with the Good Friday oil or drinking the exorcism water for the vaccinated to live without dying from the shots. Trust in Me by believing that I can heal Covid diseases and the Covid vaccinated people.”
Jesus said: “My people, you are seeing the destruction of Hurricane Ida in Louisiana and other states. Power will be out for millions of people for about three weeks if the linesmen are able to fix the damage. There is a lot of water damage as well. These people will need donations for food, water, and shelter. Your country is being punished for your abortions and your worship of worldly things instead of Me. You will see more natural disasters for the same reasons. The fact that Ida came about the same day of the year as Katrina, is another lesson that your people did not repent even amidst all of this damage. Since your people’s sins are getting worse, then you will continue to face My justice. Pray for the conversion of sinners, and pray that your leaders do not force the Covid vaccines on everyone. You need to have three months of food for each household member to face the coming food shortage or famine all over your country. If your evil leaders try to force the unvaccinated people to get the Covid shots, and threaten your lives, then I will bring My justice against them and their lives will be threatened. I will bring you to My refuges if they threaten your lives with the mark of the beast. I will make you invisible so they cannot kill you. Trust in My protection now and throughout the tribulation.”
(*) Note By a soul. Last week, before these two messages on Puerto Rico were given by the Lord, I spoke with the wife of John Leary. She told me how people who were foreign travelers had visited Puerto Rico and they were denied groceries and gasoline because they were unvaccinated. So, I believe this incident is what the Lord was referring to when He indicated twice that the unvaccinated cannot buy groceries or gasoline in Puerto Rico. I do not know anything further about the lockdown situation in Puerto Rico. Thank you. God bless!
Jesus said: “My people, you use your phones to keep in touch with one another, so in this vision of an old dial-up phone, this is My sign that I am always calling you to keep in touch with Me at Mass, in your prayers, and when you receive Me in Holy Communion. I love all of you as you are My creations, and I want to be close to you in everything you do for Me. My son, I know how much you love Psalm 27 when it speaks of how you seek to be with Me in heaven. You truly will desire to gaze on My loveliness and contemplate My love in all of My creation in heaven. In the Gospel you saw how I cast out the evil spirit from a man because I am much more powerful than the evil spirits. They are like little flies that I command to leave. The demon knew that I am the Holy One of God, and he was fearful of Me. So have no fear of the evil ones who may seem to have authority in this world, because I will soon cast all the demons and evil people into hell. You will soon see My victory in the Battle of Armageddon, and I will bring My judgment on the earth with My Comet of Chastisement that will fall on the earth and the demons and evil people will be cleansed off the face of the earth into hell. Then I will renew the face of the earth and I will bring My faithful into My Era of Peace as your reward for being faithful to My service.”
Monday, August 30, 2021
Jesus said: “My son, I told you a prophet is not accepted in his hometown. I have given you a message about the end times to share My words with the people so they will be prepared when I will bring My Warning. The best preparation for the Warning is to come to frequent Confession so you can keep your soul pure and ready to meet Me at your mini-judgment. People will come to Me outside of your body and outside of time, and you will have a life review of your good and bad deeds. You will be focused on your unforgiven sins. By your life’s actions you will receive a judgment to heaven, purgatory, or hell and you will have a taste of your present destination. The glory of My Warning will be that you will be given a second chance when you are brought back into your body and back into time, so you can change your life for the better. You will have six weeks of conversion after your Warning experience to change your life with no evil influence. This will be your opportunity to try and convert your family members and friends to being true believers in Me, so they can receive My cross on their foreheads, so they can enter My refuges. At the end of the six weeks you are to get rid of all of your internet devices so you do not look at the Antichrist’s eyes. I will soon call you to My refuges of protection during the tribulation. Your guardian angel will lead you to the nearest refuge with a flame to guide you. At the end of the tribulation, I will bring My Comet of Chastisement, and all of the demons and evil people will be cleansed from the earth into hell. I will then lift My faithful into the air, and I will renew the face of the earth. Then I will bring My faithful down into My Era of Peace.” (Thess 4:13-18)
Jesus said: “My people, you know that I go before you to prepare a place for you in heaven for the worthy souls. Your priest prayed a votive Mass for a happy death. This reminds you of St. Joseph who is a patron for the dying. When you see someone dying, you need to ask a priest to give that person the sacrament of the sick. This and Confession is a great help for a dying soul. You can also pray the Divine Mercy Chaplet for such a soul to help save this person from hell. Every person is appointed to die because your body is mortal, but your soul is immortal. You do not know when I will call you home, so you need to keep a pure soul with frequent Confession so you are ready to meet Me at your judgment at any time. There are some souls in your family or among your friends that have not accepted Me into their lives, or they are lukewarm in loving Me. By your persistent prayers and fasting for these souls, you can assist them to avoid hell, because I can answer your many prayers. Keep close to Me in your prayers every day in case you should die this day. I love all of My faithful people and those people, who wear My Blessed Mother’s brown scapular, and lead a good life, will avoid the flames of hell. Trust in Me to have My angels guard over My faithful souls, even in death.”
Sunday, August 29, 2021
Jesus said: “My people, you are seeing in the Gospel how the Pharisees followed their Mosaic practices, and they were critical of My disciples for not washing their hands before eating. Your priest made a good mention when he compared the smaller traditions to the major practices of the Church. He was asking you to think deeper in everything you do, and you should do things out of love and respect for Me. You were pleasantly surprised that the priest gave you Holy Communion on the tongue, which is out of respect for My Host that you do not touch the Host with your hands. I appreciate My faithful when they give respect to My Blessed Sacrament. You can even make time for Me in Adoration of My Eucharist. You are showing your love for Me in receiving Me worthily with a clean soul free of mortal sin. I am also happy for all of My people who come to Me at Sunday Mass. This is a fulfillment of My Third Commandment that you give Me proper worship on Sunday. Pray for your family members that they can see your example to attend Sunday Mass and monthly Confession. There are many lukewarm Catholics who need to show Me their love, so continue to pray for your family members that they will wake up during the six weeks of conversion after the Warning, or they could be lost in hell. The harder you pray and fast for them, and the more consistent that you pray for them, I will answer your prayers and they can be saved as true believers.”
Saturday, August 28, 2021 (St. Augustine)
Jesus said: “My people, you are familiar with the Parable of the talents of gold where one servant was given five talents, and he gained five more. A second servant was given two talents, and he gained two more. The third servant was given one talent, but he buried it in the ground. The Master was pleased with the first two servants and he gave them more responsibilities. The third servant had his one talent taken away and he was severely punished for wasting his time and the money given to him. The lesson is that you are given many talents to achieve your given mission, but unless your heart is open to loving Me and serving Me, it will be difficult to complete your given mission. Let your candle burn for My service as long as you live. By your service and good deeds, I will judge your life at your judgment. So do not waste your time with frivolous earthly pursuits, but keep focused on Me and carry out the directions I give you to accomplish the mission I have given you. These people, who are given more talents, will be expected to accomplish more, and they will have a harsher judgment. By following Me in all of your duties, I will say: ‘Well done, My good and faithful servant. Enter into your Master’s joy in heaven.”
Jesus said: “My people, the one world people know that those people, who receive two or more shots of the Covid vaccine, could die within a few years. This is why they are carefully recording those people who received the shots. The evil ones are even bribing people with money to take the shots. They are doing everything to force employees and students to take the Covid shots. They know the people who are resisting to take the Covid shots. There will come a time to try and collect the people who are not taking the shots. This will endanger the lives of My faithful, and I will call these people to the safety of My refuges. It is the door to door campaign to collect resisters, when I will make My people invisible so the evil ones do not see you. If more serious searches are brought, then I will bring My Warning, and call My faithful to My refuges. Fear not these gestapo tactics to try and kill the unvaccinated people, because I will protect My faithful from the evil ones at My refuges.”
Friday, August 27, 2021 (St. Monica)
Jesus said: “My people, STAY AWAKE, for you do not know the day, nor the hour that I will come to you at your judgment. This story of the five wise virgins and the five foolish virgins is My best warning to My people to be prepared for the coming tribulation. Keep your souls pure with frequent Confession, Mass, and Holy Communion. You also need to have a backpack ready with some food, water, clothes, and hygiene items. Have a small tent and a sleeping bag ready to put in your vehicle when you are called to leave your homes for My refuges. My refuge builders need to consecrate their refuge, and have a water source as a well or barrels of water. Have some food ready so I can multiply it. Have some beds, blankets, and pillows ready for sleeping. You will need some fuels to heat water, and to heat and cool the house. Some solar panels would be extra, but helpful. You will need lights at night, and pots, pans, silverware, and plates for eating. I will bring you daily Holy Communion from a priest or My angels. Have vestments, books, candles, bread, and wine for Mass if you have a priest. You will need an altar and a tabernacle with a monstrance for your Perpetual Adoration. These things are your refuge preparations that you will need for surviving, and My angels will protect you from the evil ones. Trust in Me to protect you and provide for your needs during the tribulation.”
Jesus said: “My people, you need to obey your bishop and cardinal, as long as they are directing you according to what the Church teaches. You also need to pray for your bishops and priests. My priest sons give you My Body and Blood at the Mass, and this is a little taste of heaven on earth. They also give you My sanctifying grace when they heal your soul with Confession. The priests also administer the other sacraments as well. You need to support your bishops and priests with your donations to help support My Church. Having bishops and priests are a gift from Me, as they represent Me in their office. Pray for vocations to the priesthood and for deacons who are needed to serve My people.”
Thursday, August 26, 2021
Jesus said: “My son, I have given you a previous message that you will be living into the tribulation time of the Antichrist. You will continue to have messages even at your refuge. In the vision you saw many steps to heaven because you will be living through the end times, and even into My Era of Peace. During the tribulation you will be traveling around to various refuges to support them by bi-location. People need to come to frequent Confession to keep their souls holy for the trials that are to come. Have no fear of the evil ones who want to kill My faithful, because I will call you to the safe havens where My angels will shield you by being invisible. My Warning will be coming soon due to the coming threats on your lives. After your life review and mini-judgment, you will have six weeks of conversion without any evil distractions. This is when you will have an opportunity to try and convert the weak people in your families, so they can be firm believers and they can enter My refuges. My Warning will give every sinner one last chance to convert to being a true believer. Those people, who refuse My love, and refuse to serve Me, could be choosing the broad road to hell, if they do not change. Keep praying for the souls of your family and friends, so they can be saved.”
Jesus said: “My people, you need to pray for your farmers that they have enough water to grow their crops, and that fields remain dry so they can harvest their crops. In various parts of the world food is getting scarce and fresh water is also scarce. Pray that you do not have another shutdown that could be problems for stocking your stores and that your shelves do not go empty. This is why you should have three months of food for each member of your household in case the stores go empty. Trust in Me to provide what you need to eat.”
Jesus said: “My people, the evacuation of Americans and Afghans in Afghanistan has been complicated by a suicide bomber exploding a bomb that killed some American soldiers and wounded others. Your soldiers were trying to evacuate the most Americans as possible. This slowed down the evacuation. The last planes out could see problems as the Taliban is blocking the roads. Pray that your people can be evacuated without any more incidents.”
Jesus said: “My people, you will see the Taliban persecuting the religious people as they will bring back Shariah law. Already Iran is trying to sell oil to these people in Afghanistan. This evacuation of Americans has been a disaster, and many will be trapped without any follow up support. The Democrats are being criticized for their poor planning. Pray that there will be no more killing in this evacuation.”
Jesus said: “My people, many of My believers will be shocked how quickly your country will be taken over by the Democrat communists. You are seeing vaccine mandates being forced on all parts of your people, and there will be a backlash against this control over your people’s health. If enough of your people rise up, you could see a civil war that could kill a lot of people. Your people are only quietly protesting now, but will they be willing to defend their freedoms when any violence breaks out? I will call My people to My refuges when your lives are in danger. Be ready to leave your homes for a refuge within twenty minutes with your backpack.”
Jesus said: “My people, as your schools open in person, you could see more sickness among the young children in coming back to class. Pray for your children to remain healthy. If you see a quick increase in Covid deaths and cases, you could see another possible shutdown. This could make a food shortage even worse among your people. If needed I will multiply your food so you will be able to survive a famine. Trust in My angels to prepare you to come to My refuges.”
Jesus said: “My people, at one protest on Monday you had a better organization of over 200 people. It makes a big difference when you have a means of getting the word out in good communication. Today’s protest was only about 50 people, and there was not as good a communication of time. You can see that you need numbers for a good protest. A good means of communication is also necessary so that your people know where and when to meet. Your people have a strong desire to defend your freedoms, but your numbers are few when you consider what is at stake. It is hard to educate your people about the true death dangers of these poisonous vaccines when the media and Biden are brain washing the people to take the vaccines. Pray that you can bless the vaccinated people with the Good Friday oil and the exorcism water. I will soon call My people to My refuges after the Warning, so the believers can be healed at My refuges.”
Jesus said: “My people, you can prepare spiritually for the Warning by coming to frequent Confession. You can prepare physically for the coming famine by having three months of food for every household member. All of the people will need to make a free will decision after the Warning whether they want to be with Me or with the devil. My believers will be protected by My angels at My refuges during the tribulation. The non-believers will suffer from the plagues and the Antichrist as they will be on the road to hell. Come to Me, My people, and I and My angels will help you through this time of tribulation.”
Wednesday, August 25, 2021
Jesus said: “My people, this strange vision of dirt and ashes is a sign of My justice, as I will destroy the wicked people who are trying to kill My people with their evil poisonous vaccines and viruses. Those evil politicians and doctors, who are lying to the people about the evil plan to reduce the population, will be burned and destroyed for their evil deeds. These evil ones are trying to use employers and colleges to mandate the poisonous shots on the employees and the students. Your people need to wake up and realize that these mandated shots will kill you in a few years. You are really choosing to die to keep a job when you will not be there. This is why purposely killing a lot of people is so evil, and your lying leaders and health people should be put in jail for purposely trying to reduce your population. You will soon be seeing many dead bodies on the ground from these evil shots. Shout from the rooftops not to take these toxic shots, and try to wake up your people to fight these illegal and evil mandates that should not be followed. If you protest hard enough, you may be able to stop these illegal mandates. You can use the Good Friday oil or the exorcism water to bless the vaccinated people so they will not die. In any case you will see the bodies of these evil ones burned to a crisp for their evil actions. My justice will be carried out on the evil ones who are causing these mandates. My believers will be called to My refuges of protection to avoid taking the vaccines, where you will be fed and protected from the UN troops who will be trying to kill you.”
Jesus said: “My people, I am showing you an empty donation basket. The meaning is all about how much of your time, talent, and money that you are willing to give up and contribute to saving souls. I call all of My people to work on converting souls, but this is not easy to do for people. It is the intention in your heart to serve Me that I am observing. You need to be faithful to your daily prayers and Adoration when you make time for Me. This is how you show your love for Me and your neighbor. Just as I ask you to give generously with your donation of money, so you also need to be generous with your time. You and your wife spend time giving out bottles of Good Friday oil, and you give of your time in coming to talks, and sharing My messages in your Zoom conferences. However, people can share their faith with others even in small ways every day by their actions. Coming to daily Mass is another way to witness your love for Me. I love all of you and I thank you for all of your evangelization efforts.”